THE PSYCHOPHYSIOLOGY OF REM SLEEP IN RELATION TO MECHANISMS OF PSYCHIATRIC DISORDERSVadim S. Rotenberg E-mail: vadir@post.tau.ac.il A THEORETICAL BACKGROUND - SEARCH ACTIVITY CONCEPT Numerous investigations on sleep psychophysiology in psychiatry have provided science with interesting empirical data. However, there are many contradictions in these data and the most important and substantial questions are still far from being solved. The only way to overcome these contradictions and to elucidate the role of sleep alteration in the general pathogenesis of mental disorders is to develop an integrative theory of the basic sleep functions, which are equally relevant for humans and animals, for psychology and physiology, for mental health and for psychopathology. Obviously, this is a difficult task. It would be too optimistic to expect that such an integrative theory would appear at once in its final and comprehensive version. Thus, it is worth producing different versions of such a theory for broad discussion, to provide in the future a holistic picture from all of these puzzles. The task of the present chapter is to use one such theory (search activity concept) for the revision of some contradictions and for the discussion of some theoretical assumptions. The search activity concept 1-5 deals mostly with the function of rapid eye movement (REM) sleep, although the alteration of NREM sleep seems also to be very important for understanding the mechanisms of mental disorders. However, NREM sleep deficiency is an unambiguous and general finding in many psychiatric patients with different diagnoses, while data on REM sleep are much more contradictory. The discussion of the nature of such contradictions may be very productive. Search activity is defined as "activity that is oriented to changing the situation (or at least the subject's attitude to it) in the absence of the precise prediction of the outcome of such activity but taking into consideration the results at each stage of the activity". According to this definition, search activity is a component of many different forms of behavior: self-stimulation in animals, creative behavior in humans, as well as exploratory and active defense (fight/flight) behavior in all species. In all these forms of activity the probability forecast of the outcome is indefinite, but there is a feedback between the behavior and its outcome enabling the subject to correct his behavior according to the outcome. Many forms of behavior do not include search activity. Renunciation of search is opposite to search activity and encompasses depression and neurotic anxiety in humans, freezing in animals and learned helplessness in both species. Neurotic anxiety, according to psychodynamic conceptions, is a consequence of the repression of ah unacceptable motive from consciousness. The repression can be regarded as a purely human variant of renunciation of search, of the modes of realization of the unacceptable motive in behavior, and modes of integration of this motive with other, realized, behavioral orientations. The difference between productive emotional tension (the normal anxiety of a healthy individual in a state of stress), which is instrumental in mobilizing all psychological and physical resources to overcome obstacles, and unproductive emotional tension, which hampers successful activity, is determined by the presence or absence of search activity in the structure of emotional tension6. Panic reaction can imitate search activity, but a subject in a state of panic does not consider the real outcome of his behavior and is unable to correct it. The common clinical condition of panic is depression. Stereotyped behavior is based on the precise prediction of the outcome and is not flexible; thus it also does not fit with the idea of search activity. I have proposed this new classification of behavior because our investigations have shown that search activity has a very important psychobiological meaning 1;6-13. All forms of behavior that include search activity, but differ in other respects, increase body resistance, while absence of search decreases body resistance and predisposes a person to mental and psychosomatic disorders. Persons with psychosomatic illness have a more pronounced tendency to give up than do healthy subjects, especially in stressful conditions and after facing failure. We concluded that the process of search activity in itself, independent of the pragmatic results of such activity (i.e. whether it is successful or not) and independent of the accompanying emotions, protects persons from somatic disorders. Herein lies the basic difference between the concept of search activity and the concept of coping behavior 14 that must be successful in order to be protective. The protection of search behavior reflects a particular self-service and a positive feedback: search activity requires a lot of effort and stimulates a person to enter unpredictable and potentially dangerous situations, and prevents exhaustion in such situations. All details of the investigations performed on animals and humans as well as all aspects of the theoretical discussion can be found in the above-mentioned publications. The biochemical mechanisms of search activity have been insufficiently studied but it is possible to suggest that the brain's monoamine system is closely connected with search behavior. Learned helplessness accompanied by somatic disturbances emerges when the brain monoamine level drops 15. An artificial reduction of brain catecholamine levels by tetrabenazine speeds passive behavior whereas the prevention of its depletion by monoamine oxidase (MAO) inhibitors raises stress resistance, restoring the animal's ability for an active reaction to stress 16. In animals that cannot control the stressful situation the brain level of norepinephrine drops particularly low and they show the greatest distress 17,18. Finally, the locus ceruleus plays a major role in the organization of various forms of search behavior 19. On the basis of the preceding facts the following hypothesis has been advanced 8-20. Search activity can begin in the presence of a certain critical level of the brain monoamines that are utilized in the course of search. Search activity itself, once it starts, stimulates the synthesis of the brain monoamines and ensures that they remain at adequate, or even excessive, levels. The task of such a system with positive feedback (brain monoamines -behavior - brain monoamines) is to support search activity. At the same time, a stable high level of brain monoamines causes a hyposensitivity of the inhibitory alpha2-adrenoreceptors. As a result, in order to switch on the negative feedback that regulates homeostasis of the brain monoamines, its level has to be excessively high. In a state of renunciation of search the above-mentioned positive feedback between brain monoamines and behavior does not function. Moreover, in this state, which manifests itself in depression and maladaptive emotional tension, "the monoamine expenditure climbs. The requirement for brain monoamine production and expenditure decreases because the behavior that requires brain monoamines is absent. At the same time, the low level of brain monoamines by itself does not stimulate its own production. The brain monoamine system is only partly homeostatic. If the appropriate behavior (search activity) is absent, activation of the brain monoamine system is not required. When the level of brain monoamines is constantly low the inhibitory oc2-adrenoreceptors become adapted to it and maintain this low level. If search activity is of such great biological relevance, and the absence of search, and especially renunciation of search in stressful conditions, leads to illness and death, why has the latter regressive behavior survived in the course of evolution and what makes it emerge? All higher mammals, including humans, at the early stages of ontogenesis inevitably experience helplessness determined by the relatively slow development of the central nervous system and all mechanisms (nervous, hormonal and vegetative) that ensure subsequent search behavior. Naturally, during the early stages of ontogenesis such a state cannot be called renunciation of search. It is normal and inevitable and the only accessible form of a defense reaction for an immature organism. Nevertheless, the organism thus acquires an early experience of passive reaction, an experience of helplessness. Both this experience and the ways to overcome it are of colossal relevance to the individual's entire subsequent life. In the case of the correct attitude of the primary group, above all the mother, this early experience of helplessness can be successfully and painlessly overcome. However, all injuries of early childhood, from physical separation from the mother to insufficient emotional contact with her and the feeling of insufficient protection as a result of strained relations between the parents, can consolidate the experience of primary helplessness and produce a tendency to give up. The imprinting mechanism may lead to the development of a state of renunciation of search in an adult, especially if the emotional problems encountered in some way resemble the conflicts of early childhood. Thus, readiness for the development of mental and psychosomatic illnesses is formed. It is exactly from these general biological positions, the relations between early helplessness and the formation of search activity, that the present author proposes considering the Freudian theory of the role of early psycho-traumatizing situations in the entire subsequent development of the individual. Regressive behavior in the case of neuroses and psychosomatic illnesses, according to Freud, is indeed a regression towards a biologically earlier state of helplessness that assumes a form of renunciation of search. However, if search activity is so important for survival, and renunciation of search is so destructive and dangerous for health, it would be reasonable to assume a natural brain mechanism that can restore search activity after temporary renunciation of search. I believe that dreams in REM sleep fulfill this function. A covert search activity in dreams' compensates for the renunciation of search in previous wakefulness and ensures the resumption of search activity in subsequent wakeful-ness. I base this claim on the following findings: (1) Different forms of animal behavior that contain search activity (self-stimulation, fighting) suppress REM sleep in subsequent sleep without a restorative rebound effect 21-22. This means that less REM sleep is required after such overt behavior. (2) Renunciation of search produced by the direct stimulation of the hypotha-lamus causes an increase in REM in subsequent sleep 1. This corresponds to data 23 that REM sleep increases along with a moderate reduction of norepinephrine system activity, while the excessive decrease or increase of this activity suppresses REM sleep (see discussion in references 12, 20). This means also that search activity can start in REMS sleep with a lower level of brain monoamines than is required in wakefulness. This conclusion corresponds to data 25 that in REM sleep alpha2-adrenoreceptors are much more sensitive than in wakefulness; thus the fine regulation of brain monoamines can be performed on their lower level. (3) If, during REM deprivation, the subject is involved in active behavior (exploration, active defense reaction), the REM rebound effect during subsequent sleep is substantially reduced 26. (4) Depression, neurotic anxiety and generalized learned helplessness as manifestations of renunciation of search are accompanied by increased REM sleep requirement (REM sleep latency is decreased while the duration of the initial REM episodes is increased) 27-29. A correlation was detected between learned helplessness and REM sleep percentage 30-31. (5) Both REM sleep and search activity in wakefulness are characterized by a regular and synchronized hippocampal theta-rhythm. Moreover, the more pronounced the theta-rhythm in wakefulness, the less pronounced it is in subsequent REM sleep 26. REM sleep in animals regularly contains pontogeniculo-occipital (PGO) spikes, which are electroencephalographic (EEG) signs of neural activation. In wakefulness, PGO correlates with orienting activity 32. The presence of the PGO spikes in REM sleep may mean that the animal is predisposed to react to novel stimuli, including spontaneous changes of dream content. (6) When a special part of the nucleus ceruleus (nucleus ceruleus aleph) in the brainstem is artificially destroyed and as a result muscle tone does not drop during REM sleep, animals demonstrate complicated behavior that can be generally described as orienting (or search) activity 33. (7) The same psychological variables that predispose the subject to renunciation of search - trait anxiety, fixation on obstacles according to Rosenzweig 34, motivation of avoidance of failures - have a tendency to correlate with REM percentage 8. (8) REM sleep is increased in students who display a stable emotional tension both before and after an examination, even with its positive outcome (when the stress situation no longer exists) as compared with students who display emotional tension only before an examination 2. It is possible to suggest that this stable emotional tension, which is not compatible with the actual situation, is similar to neurotic anxiety and represents renunciation of search. (9) Healthy subjects with normal search activity during wakefulness are characterized by active participation in their own dreams. This active participation correlates with heart rate acceleration and eye movement density in REM sleep 35. (10) The more characters and descriptive elements that appear in the dream, and the more active the characters and the dreamer himself, the larger the decrease in the scale of unhappiness during the night 36. It can be suggested that the characters' activity represents search activity in dreams. One must stress that, in the case of healthy subjects, the decrease of unhappiness after sleep correlates only with the above-mentioned dream variables, not with the sleep physiology. As I see it, this means that, as long as the dream is functionally effective and is able to restore search activity, REM sleep requirement does not increase. It is increased only when dreams are going to lose their restorative capacity, for instance in clinical or pre-clinical disorders (depression and subsyndromal depression, neurotic anxiety, narcolepsy). The dreams of healthy subjects represent a very specific kind of search activity, which, however, is compatible with the above-mentioned notion of search activity: the subject is active in his dreams but is unable to make a definite forecast according to the outcome of dream events. The dream does not apparently fit solely those parts of the notion emphasizing the permanent consideration of the behavioral outcome. This is true only if the dream is not self-reflective and does not include self-control, i.e. if activity in the dream is a chaotic activity. However, it was demonstrated 37 that, in the vast majority of spontaneous dreams, the dreamer is moderately self-reflective and effective in dream control. Dreams provide a good opportunity for search activity after giving up. First, the subject is separated from reality while sleeping, including those aspects of reality that caused renunciation of search. Thus, in dreams a person is free to start from the beginning. Second, within his dream, the dreamer can be very flexible in his behavior: it may be an attempt to solve an actual problem in a metaphoric manner, or it can be an attempt to solve another problem, one that displaces the actual problem 38, since not the topic of search, but the search process itself is the main restorative factor. Moreover, in dreams, one is not restricted by logical and conventional rules while manipulating the problems. One is able to use image thinking, which is polysemantic by nature and as a result much more flexible than logical thinking and, unlike the waking consciousness, can avoid contradictions 5,20,24,39,40. Image thinking is free from the probability forecast 41,42. Since we assume that the final aim of the dream work is not the real solution of the actual problem but only the restoration of search activity, all the above features contribute substantially to the restoration. The proposed explanation of the functional significance of the REM sleep in the restoration of search activity helps us to understand a very well-known phenomenon in human sleep, the "first-night effect" (FNE). It is an alteration of sleep structure observed on the first night of the multinight sleep study relative to subsequent nights 43,44. Compared with later nights, the first night in the laboratory is characterized by increased REM sleep latency, combined with a moderate REM sleep reduction, especially in the first part of the night, usually without the subsequent REM sleep rebound. Sleep latency is also often increased and sleep stages shift, while sleep efficiency is decreased. FNE reflects the natural increase of the sleeper's state of vigilance in an unfamiliar sleeping environment as part of the adaptive orientation in the new situation 45, and is much less prominent in healthy subjects sleeping at home where they are reacting only to the monitoring equipment 44,46. Such adaptive orientation includes search activity (orienting behavior, as I have stressed, is a manifestation of search activity); thus according to the search activity concept, the requirement of REM sleep in the night sleep has to be decreased. That is exactly the case. It means that the FNE can be used for the estimation (even quantitative estimation) of the subject's ability to display search activity. Another theoretical outcome of the search activity concept is the conclusion that REM, sleep may be efficient or inefficient according to its main function - restoration of search activity 5,35. If dreams do not contain a covert search activity, and moreover, if it is a continuation of the renunciation of search in dreams, it is possible to suggest REM sleep as inefficient. REM sleep inefficiency displays itself not only in the stable feeling of helplessness and giving up in dreams, in the passive position of the dreamer facing the dangerous events in the dream, but also in the decrease of dream reports after awakenings in REM sleep and in the impoverishment of these reports (fewer images and events). Such features characterize, for instance, dreams of depressed patients 38,47,48. As mentioned before, there are some physiological correlations of the subjects' imaginative activity in their own dreams. Thus, it is possible to use such physiological variables for the estimation of REM sleep functional activity even without awakenings. In the following sections of this chapter I will discuss alterations of sleep psychophysiology in different mental disorders using the frame of the search theory.
POST-TRAUMATIC STRESS DISORDERS
Complaints about sleep disorders - difficulty falling or staying asleep, increased number of spontaneous awakenings, recurrent distressing dreams (nightmares) - belong to the main diagnostic features of post-traumatic stress disorders (PTSDs). In most cases these complaints are confirmed by polysomnography: the sleep of PTSD patients is characterized by disturbances in sleep continuity, including increased sleep latency, increased number of awakenings, especially awakenings in REM sleep during nightmares, and low sleep efficiency 49-54. However, data on sleep structure in PTSDs, and especially data on REM sleep variables, are very ambiguous and contradictory. First, PTSDs are characterized by a high variability of REM sleep latency. In comparison to healthy control subjects, REM latency in PTSD patients has been reported to be either shortened or lengthened 55-60. Correspondingly, REM sleep time was either increased 61 or decreased 57. In some investigations, there was no difference in the mean REM time and REM latency between healthy subjects and combat veterans 62. PTSDs are often accompanied by major depression (MD) or other depressive disorders such as dysthymia 54, 63, 64. Thus, it has been reasonable to assume that the difference between PTSD patients in REM sleep variables may be determined by the presence pr absence of PTSD co-morbidity with depression 54. Comparison of PTSD and MD patients has shown that REM sleep time was reduced in PTSDs; however, this reduction was presumably a side-effect of the decreased sleep duration 54. In contrast to healthy subjects, the decrease of the total sleep time in PTSD patients was not accompanied by a compensatory increase in the percentage of REM and slow-wave sleep (SWS). At the same time, REM latency does not differ significantly between both clinical groups. However, it was difficult to draw a definite conclusion from this study that PTSD patients display REM sleep latency typical for depression. First, depression in the group of MD patients was not severe, and only 47% of these patients had REM latency less than 60 min. On the other hand, 20% of PTSD patients suffered from co-morbid MD, and 40% of these patients had prior depressive episodes. Moreover, within the PTSD group there was no significant trend for REM latency to be reduced in patients with the current co-morbid depression. Thus, the suggestion of depression as a reason for the high variability of REM latency in PTSDs was neither confirmed nor refuted. The investigation of Woodward and colleagues 65 seems to give a more definite answer to this question. Two groups of PTSD patients - one with and one without co-morbid depression - demonstrated similar mean REM sleep latencies, although the variability of REM latency was higher in patients with a co-morbid depression. Thus it is not depression itself, or at least not only depression, which determines the decrease of REM latency in some PTSD patients and causes heterogeneity of REM latency in all groups of these patients. The authors concluded that "a (unknown) factor emerges in PTSD which exerts a specific effect upon REM sleep timing and amount. ... This factor is related to the apparent increase in both tonic and phasic REM phenomena in sleep of PTSD patients". In discussing this topic, it is also worth emphasizing that some core physiological features of PTSD are the converse of those in depression. PTSD patients in comparison to healthy subjects, and contrary to depressed patients, are characterized by the decreased basal cortisol level and by the increased negative feedback regulation of cortisol excretion (see reference 66). Patients with PTSDs demonstrate augmented cortisol suppression in response to dexamethasone, while depressed patients are characterized by non-suppression of cortisol in this test 67,68. PTSD patients have more, and depressed patients have fewer, type II glucocorticoid receptors in circulating lymphocytes than do normal controls 69,70. Platelet alpha2-adrenergic receptor binding, platelet MAO activity and platelet serotonin uptake are decreased in PTSD patients in comparison to normal subjects 71. The increased activity and reactivity of the monoamine system in PTSDs is confirmed by increased nocturnal noradrenergic excretion 53. Thus, it is possible to suggest an exaggeration of the normal adaptive physiological response to stress in PTSD - although without any real stressful condition - and this exaggerated response to stress is the opposite to what may be found in depression. This exaggeration of the normal response is confirmed in the investigation of the FNE 45. According to the REM sleep variables, and especially to REM latency, PTSD inpatients were bidirectionally sensitive to the degree of familiarity they associated with the sleep laboratory conditions. In comparison to non-ill trauma-free controls, in-patients familiar with the laboratory environment demonstrated reduced FNE (increased extinction of the orienting reaction), while patients unfamiliar with this environment demonstrated enhanced FNE. At the same time MD patients, in contrast to normal subjects, often demonstrate a reduced FNE in the unfamiliar situation (see below). Thus, PTSD symptomatology is very complicated. On the one hand, depression is a common PTSD co-morbidity and there are some factors that make REM sleep variables in isolated PTSD similar to those in depression. However, on the other hand, PTSDs differ from depression according to many physiological variables. We will show that this distinction has real roots in the clinical picture and in the pathogenesis of PTSDs. The clinical picture of PTSDs includes, on the one hand, a persistent re-experiencing of the past traumatic events manifested as intrusive distressing recollections (images, thoughts, perceptions, dreams, dissociations like illusions, hallucinations and flashback episodes) accompanied by increased physiological reactivity on exposure to internal or external cues that symbolize or resemble an aspect of the traumatic event. It can be assumed that persistent symptoms of increased arousal such as irritability, hypervigilance, exaggerated startle reaction, prominent orienting reaction and FNE, difficulty falling and staying asleep, although they are usually considered separately, belong to the same group of symptoms - group I. On the other hand, the same patients (and this is worth emphasizing) display persistent avoidance of stimuli associated with the trauma and numbing of general responsive-ness (conscious avoidance and repression of thoughts, feelings, memories and activities associated with the trauma; diminished interest in any forms of activity and interpersonal relationships, as in depression; a hopeless view on the future). This group II of symptoms is obviously opposite to group I, and both groups compose a contradictory clinical picture, where symptoms of group I are continuously displaced by symptoms of group II and vice versa, or even partly appear together. However, it is possible to integrate these different symptoms in one holistic frame. The central point of the PTSD diagnosis is the feeling of helplessness caused by the traumatic event. It is the central feature of the entire clinical picture. Fear of death and fear of ego-destruction produces helplessness because it is a very distressing and disorganizing emotion - there are no clear ways to cope with a fear of death, if the psychological defense mechanisms become insufficient 72. The feeling of helplessness is very difficult to resist. It is a state of giving up, of renunciation of search, with all negative outcomes on human behavior and health, as shown in the Introduction. By considering helplessness as a corner-stone of PTSDs, it is possible to explain all group I symptoms as attempts to cope with this feeling of helplessness and loss of control which accompanied the traumatic event. Hypervigilance, and exaggerated startle reaction, reflect the mobilization for coping. However, unfortunately, this coping is irrelevant and cannot be successful, because the object of coping is elusive: it is the traumatic event that already happened in the past, and the subject is unable to win back. In principle, the re-experience of trauma may help to re-examine the traumatic event and to include it in a more broad and polydimensional picture of the world where trauma is able to find its definite and restricted place and will no more cause general helplessness. However, for this re-examination, the subject has to be well equipped with skills of the right-hemispheric polysemantic way of thinking 39,73, and exactly these skills are underdeveloped in PTSD patients 74. In these patients traumatic experiences are fragmental, not integrated in the holistic picture of the world, and reappear in a very rigid form of primitive and frustrating images in wakefulness, in altered states of consciousness and in nightmares. Such coping is stereotypic in its nature; however, it can include a very strong physiological mobilization. The position of the subject in this state of coping can be very active, but the final goal - a freedom from helplessness and from a feeling of being a victim - cannot be achieved. It is like a fight with a shadow where you never can win. Traumatic events, instead of being coped with, restore every time the same distressing frame and do not help to overcome helplessness. It is a key point of this unsuccessful coping. Group II symptoms reflect an attempt to avoid the negative experience with which the subject is unable to cope. To avoid does not mean to solve, but only to conserve the problem. Trauma after such avoidance remains isolated from the normal integrative function of the brain and from the larger associative network of memory. Actually, these group II symptoms display a giving-up state in front of traumatic experience after the unsuccessful coping. However, giving up only increases helplessness, and it forms a vicious circle. Coping and avoidance work in opposite ways towards the same .goals, but both are unsuccessful. The continuous oscillations between unsuccessful attempts to cope and giving up (avoiding) can explain the complicated and contradictory picture of PTSDs and their sleep alterations. For instance, Woodward and co-workers45 proposed that helplessness and not clinical depression may be a factor related to increases in both tonic and phasic REM phenomena and reduced REM latency. Conversely, the predominance of group I symptoms (active coping) may be responsible for increased REM latency in some PTSD patients and in the exaggeration of FNE. The same patient may demonstrate different sleep structures at different time points. By taking into consideration the role of REM sleep and dreams in balancing behavioral attitudes and restoration of search activity, it is reasonable to pay attention to the dream content. In contrast to the increased variability of REM latency and REM sleep time, eye movement (EM) density in REM sleep in PTSD patients seems to be unambiguously increased 54,62,66. According to the above-mentioned relationships between EM density and dream content 35,75, it is possible to suppose that the increase of EM density in PTSD patients correspo'ficts to their typical complaints of traumatic dreams and nightmares 59,76-79. However, direct investigation of this topic in the sleep laboratory does not confirm this proposition. According to Greenberg and associates 55, most REM awakenings in Vietnam War veterans with PTSD symptoms led to content-less reports. Kramer and colleagues 81 have also shown a lower than normal (around 50%) dream recall rate after spontaneous awakenings from REM sleep. In the investigation of the Lavie group 49,82 these findings have been substantially enlarged, and they deserve elaboration. The authors systematically collected dreams from REM sleep in well-adjusted and poorly adjusted Holocaust survivors, and compared them to the control group of aged persons without the Holocaust experience. The estimation of the level of adjustment was based on clinical interview regarding all main areas of life: marital and familial problems, problems at work, social relationships, somatic and mental problems, and general satisfaction in life. It is worth stressing that the well-adjusted group presented not only fewer somatic complaints man the poorly adjusted group, but also even fewer complaints than the control group. Members of the well-adjusted group were characterized by stronger ego-forces, in comparison to the poorly adjusted group. Their emotional feelings were positive. Recollection of the Holocaust contents occurred infrequently and was under their control. The less well-adjusted group was disturbed by their Holocaust experience. According to the clinical picture, the less-adjusted group displayed a moderate PTSD complex, including decreased sleep efficiency in comparison to both other groups. In sleep structure, including REM sleep variables, no differences were found among the groups. (This is not very surprising if we consider the variability of sleep in PTSD patients.) The most prominent differences between groups were in dream reports and in dream content. The control group reported a normal percentage (80%) of dream content when awakened from REM sleep. The percentage of reports in the less well-adjusted group was 50.7% - equal to the percentage reported by the PTSD patients of Kramer and associates 81, and Bleich and co-workers 83. This percentage is also approximately equal to the percentage of dream recall (54.6%) presented by mentally ill patients (with depression, anxiety, hypochondria and hysterical neurotic disorders 84). At the same time, the well-adjusted group had the lowest dream rate (33.7%). Moreover, in contrast to both other groups, well-adjusted survivors were unable to realize not only the content of the dream, but also even the fact of dreaming. The survivors' dreams, particularly those of the well-adjusted group, were less complex and salient in comparison to dreams of the control group. The number of anxious dreams was high in both groups of survivors and especially high in the poorly adjusted group. However, only the less well-adjusted survivors experienced nightmares resulting in spontaneous awakenings. In dreams of the less-adjusted group the dreamers were usually the victims, while in the well-adjusted group dreams were characterized by hidden hostility in which survivors were not directly involved. This is a very important point, because according to the search activity concept a feeling of being a victim means the continuation of giving up and the renunciation of search from wakefulness into dream, and this feeling reflects the functional insufficiency of the dream. Only the well-adjusted survivors demonstrated indifferent responses toward their dream content after awakening, even if the content was stressful. Similar to controls, and in contrast to the less well-adjusted survivors, the well-adjusted subjects dreamed about their present life rather than about the past. Dreams of both groups of survivors contained more danger to the dreamer, more anxiety and hostility, and more interpersonal conflicts than dreams of the control group, although all these features were more prominent in the less-adjusted group. By taking into consideration this similarity, and the relative similarity in the low number of dream reports, Kaminer and Lavie 49 concluded that both groups, and especially the well-adjusted one, were characterized by the repression of dreams, and the stronger the repression, the better was the psychological adjustment. However, this explanation looks very doubtful. First, it has already been stressed that according to many important features (such as general physical and mental health, sleep efficiency, psychological response to the negative dream content during sleep and after awakening, position in dream - victim versus non- victim) the well-adjusted group of survivors is closer to the healthy control group than to the poorly adjusted group, while according to the number of dream reports it is most different from the control group. Second, it is very difficult to accept the statement that repression is a healthy defense mechanism. As has been shown previously, repression is a very specific human form of renunciation of search, according to the psychodynamic concept of neurotic free-floating -anxiety. We have shown 85 that in psychologically maladapted somatic and psychosomatic patients there is a positive correlation between repression (measured according to Plutchik and associates 86) and the D scale of the Minnesota Multiple Personality Inventory (MMPI), which measures depression and neurotic anxiety. This correlation was absent in psychologically adapted subjects; thus it is possible to suggest that high levels of repression caused increased anxiety. If anxiety required repression for its compensation, such positive correlation would be present in psychologically adapted subjects. In addition, the high strain of repression characterized essential hypertension. This means that repression does not belong to the most adaptive defense mechanisms. Moreover, repression increases together with anxiety after REM/dream deprivation 80,87. This means that in normal subjects dreams are used for the compensation of repression and for the prevention of anxiety, and it would be very paradoxical to assume that the repression of dreams serves psychological health. It is possible to suggest repression of the functionally insufficient dreams to explain the lack of dream reports in the poorly adjusted group of PTSD patients; however, it looks very doubtful that repression can serve psychological health and is used by well-adjusted survivors. The search activity concept provides another explanation of the extremely low number of dreams in the well-adjusted group. Members of this group appear to have a very strong search activity. They are usually successful and active business people, with high social achievements 49; they cope with problematic situations well, perhaps better than people without the Holocaust experience. This resembles hyper-compensatory activity and can explain their normal health as well as a decreased dream requirement. Thus, the number of dream reports is decreased in this group not owing to the dream repression but to the high search activity during wakefulness. However, in the less-adjusted group the low number of dream reports reflects suppression of the functionally insufficient REM dreams 35. Thus, dreams in FTSDs are either anxious and hostile (coping) or repressed (avoidance of experience).
DEPRESSION
Sleep in depression has been investigated in more detail than sleep in any other psychiatric disorder and there is a general consensus regarding the sleep structure in depression. Slow-wave sleep (SWS) deficiency, especially in adults; decreased sleep duration caused by the increased sleep latency; awakenings during the night sleep and early morning awakenings; decreased REM sleep latency and the redistribution of REM sleep with its concentration in the first half of the night; increased EM density in the first cycle -various combinations of these features characterize the sleep of depressed patients 64,88-91. However, what these alterations mean and the cause-and-effect relationship between them and other clinical symptoms of depression is still an open question. There is contradiction in the literature regarding REM sleep variables: they have been claimed to normalize during remission 92,93, but this finding has not been confirmed in other investigations 94,95. Thus, the alteration in REM sleep variables may be a trait or state marker of psychopathology 96-97. REM sleep deprivation was used as an effective treatment for depression 98; thus REM sleep may even play a role in the pathogenesis of depression. Alternatively, REM sleep redistribution may reflect either an altered circadian rhythm 99 or an attempt by the brain to compensate for depression. This latter proposition appears to be more explanatory and ties in with theories concerning the adaptive function of REM sleep in healthy subjects, especially in the domain of mood 28,36,100. The search activity concept also fits this proposition because depression is a typical state of renunciation of search that requires search activity in dreams for compensation and for the restoration of overt search behavior in the subsequent wakefulness 5. According to this point of view, the decreased REM latency and increased REM duration and EM density in the first sleep cycle reflects an increased requirement in REM sleep. This increased REM sleep pressure in depression was confirmed in our recent investigation 101: while in normal subjects and in schizophrenic patients the incorporation of wakefulness in the first sleep cycle increases REM latency in proportion to the incorporated wakefulness, it does not happen in depression. The absence of the FNE, according to REM sleep variables, in any patients with severe depression 102-105 can be considered not only as an outcome of the diminished orienting reaction in depression, but also as a sign of the increased REM pressure. In our investigations 106,107, FNE was present only in 44% of patients with MD. According to our criteria, FNE was present if REM sleep latency on the first night of study was at least 30 min longer than REM sleep latency on the second night. FNE was absent in most patients with mood-congruent psychotic features (delusional depression) highly resistant to the antidepressant drugs and with more previous hospitalizations (as was shown also by Ansseau and co-workers 89), and clinical improvement in these patients without FNE was achieved only after electroconvulsive therapy (ECT). Thus, absence of the FNE characterized depressed patients in a more severe clinical state. However, and especially interesting, depressed patients who exhibited FNE also demonstrated increased REM sleep pressure. The delayed first REM sleep period in the first night was increased, in comparison to the first REM sleep period in the subsequent nights of patients with FNE as well as in comparison to the first REM sleep period in all nights of patients without FNE. In depressed patients the first REM sleep period duration is usually increased, in comparison to healthy subjects, but in patients with FNE the first REM sleep period on the first night was especially high and occupied 44% of REM sleep. Moreover, REM sleep latencies on the second and third nights in patients who exhibited FNE were significantly shorter than REM sleep latency of the corresponding nights of the group without FNE. In patients with FNE total REM sleep was increased in the second night compared with the first night. In addition, patients with FNE showed an increased number of short cycles (less than 40 min) in all nights. By taking into consideration all these data, it is possible to suggest that depressed patients who display the FNE are also characterized by the high REM sleep requirement; however, they are more flexible in their physiological responses, in particular in the response of their REM sleep system, in comparison to patients without FNE. Being in an unfamiliar environmental condition, they are able to react to this condition with a FNE, like healthy subjects - by increasing the REM sleep latency. However, presumably due to the high REM sleep requirement, they demonstrate a 'rebound effect' not typical for healthy subjects: a shift of REM sleep to the first cycle and an increase of REM sleep in the subsequent nights. I cannot exclude that they are less resistant to the treatment partly owing to this flexibility of orienting and REM sleep systems. In parallel with the increased REM sleep requirement, it could be suggested that REM sleep in depression has an adaptive function. This suggestion is partly based on the relationship between REM sleep quality and SWS restoration during night sleep. As was stressed previously, depression in general is characterized by SWS deficiency, and SWS is often restored only after successful treatment 102,108. Patients who display FNE exhibit more SWS in their night sleep 106. In addition, SWS in depression is often redistributed compared to healthy subjects: it predominates in the second, and rarely even in the later cycles. We have shown 109 that in REM sleep episodes just before such an "explosion" of SWS in the third or fourth cycle, EM density is increased in comparison to all previous REM episodes, while such an explosion does not happen in depression with a flattened EM density distribution. Thus, it is possible to speculate that in some particular cases REM sleep, characterized by enhanced EM density, contributes to the improvement of the mental state of depressed patients and determines the psycho-physiological condition that allows SWS to appear. This conclusion is in agreement with data of some recent investigations: the increase of the positive dream content from the first to the last REM sleep periods is related to the remission from depression 27; at the same time the increase of REM density from the first to the last sleep cycle in depressed patients (a relatively rare finding) is accompanied by mood improvement from evening to morning 119. However, the concept of the restorative and compensatory function of REM sleep at the first glance contradicts some experimental data, and this contradiction requires explanation. First, this concept does not correspond, to the beneficial effect of REM sleep deprivation 98 . Second, the duration of the first REM sleep period and its EM density are usually increased in depression 91; however, this does not contribute to mental health improvement. When EM density is highest in the first cycle, mood does not change from evening to morning, or even becomes worse. While EM density in the fourth cycle correlated positively with mood restoration, REM sleep duration in the first cycle correlated negatively with the same variable 110, 118. Also, the high EM density in the first REM sleep period, even if it exceeds EM density in any other cycle, does not predict or determine the increase of SWS in the second cycle. As stressed above, only in the second part of the night, after the second cycle passed, was any significant increase of SWS preceded by enhanced EM density. Previously, we have shown 2 that not the first, but the second REM sleep period increased in healthy students in the process of adaptation to stress. Thus, it is possible to suggest that the first REM sleep period is functionally different and less efficient in comparison with the subsequent REM sleep periods. According to Cartwright and colleagues 27-28, it looks as if the first REM period serves as a bridge between the emotional state of the preceding wakefulness and its regulation in the subsequent REM sleep periods. In healthy non-sensitive subjects without strong negative pre-sleep mood dream activity, the first REM period is usually short and less elaborate, and corresponds to the short REM duration and low EM density. A shift toward more intense dreams in the first REM period may represent a response to excessive levels of pre-sleep negative affect, and in such dreams this affect is still not compensated. A normal dynamic in normal subjects with a high pre-sleep negative affect is the change from the initial negative affect in the first dream to increasing positive affect in subsequent dreams. According to Cartwright and Lloyd 111, depressed patients who had an enhanced dream-like quality of mentation during the first REM sleep period showed decreased Beck Depression Inventory scores at follow-up assessment. An enhanced dream-like activity may indicate that the brain starts to deal with negative affect in the first REM period. Such dealing is a necessary but insufficient condition for adaptation and mood regulation. The sufficient condition may be the restoration of search activity in the subsequent dreams. However, when speaking about resistant depression such restoration is usually not available. There are doubts whether the normal adaptive process even starts in the first REM period, in spite of its increased duration and enhanced phasic activity. There are even more doubts about the functional sufficiency of subsequent REM periods characterized by flattened EM density, in contrast to normal subjects who display an increase of EM density from cycle to cycle 91. In depressed patients the number of dream reports after awakenings in REM sleep during the night is decreased, dreams are shorter and simpler, and contain fewer images than dreams of healthy subjects 112. We have found that in patients with MD, subjective estimation of sleep latency correlated with EM density in the first REM sleep period and subjective estimation of the number of awakenings correlated with the total EM scores 113. In healthy subjects, as has been shown previously 35,75, EM density correlates with dream reports. Thus, it is possible to suggest that in depression psychic activity in REM sleep is often not perceived as dream mentation. Dreams may be repressed from the consciousness if they contain failures and feelings of helplessness. As a result, REM sleep is considered subjectively as wakefulness. This may be one of the reasons for the underestimation of sleep duration in depression. However, at the same time it is a sign of the proposed functional insufficiency of REM sleep that it does not provide an opportunity for the restorative search activity in dreams 5,35. Thus, the increased need for REM sleep in depression is combined with the functional insufficiency of REM sleep. The insufficient dreams cannot restore search activity; moreover, such dreams may conserve or even increase the renunciation of search. The functional insufficiency of REM sleep and dreams may explain the positive outcome of REM deprivation in depression.
SCHIZOPHRENIA
In patients suffering from schizophrenia, polysomnographic findings are less consistent than in depression. Some authors noted SWS reduction 114; however, this was not confirmed by other investigators 115, 116. Lauer and associates 117 did not find any significant differences in sleep structure between a paranoid group of schizophrenic patients and a healthy control group, while, according to Mendelson and co-workers 118, REM sleep percentage increases exactly before the exacerbation of psychotic symptoms and tremendously decreases during the exacerbation. In some studies REM sleep latency was reduced 115,119-121, while in other studies it was normal 122-124. Some authors 121 suggested that symptoms of depression may be related to the decrease of REM sleep latency in schizophrenia; however, in other investigations 114, 125, REM latency was not related to depressive symptoms. The difference between schizophrenia with the predomination of positive and negative symptoms in numerous clinical and physiological variables allows a prediction that this difference may also influence sleep architecture. Some sleep investigations have been performed using this paradigm directly or indirectly 115,120,122,126. These investigations concentrated mostly on the relationships between positive versus negative symptoms, SWS and REM sleep latency. We investigated 34 patients with chronic schizophrenia on stable doses of neuroleptic medication10,127. They were divided into two groups according to the positive and negative syndrome score (PANSS) rating128. Group I contained seven men and two women (mean age 40 years) with positive/negative ratio > 1. Group II contained eight men and two women (mean age 41 years) with positive/negative ratio < 0.5. Thus, positive symptoms dominated in group I (28.1 vs. 11.8 in group II) and negative symptoms dominated in group II (30.8 vs. 20.0). The neuroleptic treatment was approximately similar in both groups. Sleep data were collected on two consecutive nights. There were no significant differences between both groups according to most sleep variables except for REM sleep percentage and EM density in REM sleep. In group I the mean REM sleep percentage was significantly lower (20%) than in group II (25%). REM sleep in the first cycle displayed a tendency to be higher in group II, but REM sleep latency was similar in both groups. We also compared positive and negative symptoms in 11 patients with more than 25% REM sleep (group A) and in 14 patients with less than 20% REM sleep (group B). The mean level of positive symptoms was 15 m group A, and 20 in group B, while the mean level of negative symptoms was 27 and 25, respectively. Negative symptoms were relatively more predominant in group A with the increased REM sleep percentage. REM sleep latency demonstrated a non-significant tendency to be shorter in the group with an increased REM sleep percentage; however, it was in the normal range. According to these data the ratio between positive and negative symptoms is not the only factor determining the representation of REM sleep. In group I this ratio was > 1; however, the mean REM sleep percentage was 20, while in group B, where REM sleep was less than 20, this ratio was less (0.8). On the other hand, in group II positive/ negative ratio was 0.38 and REM sleep percentage was 25%, while in group A, where REM percentage was more than 25%, this ratio was 0.55. In control healthy subjects matched to patients with schizophrenia according to age, the mean REM sleep percentage was 19%, very similar to patients in group I. These results suggest that a strong predominance of negative symptoms is associated with a relative increase of REM sleep percentage. However, the predominance of positive symptoms is not associated with REM sleep suppression less than the normal level and there is no linear relationship between REM sleep variables and the positive/ negative symptom ratio. Also, we have estimated the relationship between EM density (averaged EM frequency in 1 min of REM sleep) and positive versus negative symptoms. In subgroups with a high (> 6) and low (< 2.5) mean EM density the level of negative symptoms was almost equal (31 and 33), but the level of positive symptoms was significantly higher in the subgroup with low EM (25.2 vs. 12.6). This finding was confirmed in further analysis of the data. In patients with a low score of positive symptoms, mean EM density was significantly higher than in patients with high scores 127. At the same time in patients with high and low scores of the negative symptoms EM density was similar. In patients with positive/negative symptom ratio < 0.4, EM density achieved maximum values in the first cycle. In other cycles, it was lower and almost equal, thus resembling EM distribution in depressed patients129. A significant negative correlation was found between the score of the positive symptoms and EM density in the first and third cycles, -0.45 and -0.50, respectively. These findings are in agreement with Neylan and colleagues 125, who reported an increased severity of psychosis after neuroleptic withdrawal simultaneously with the decrease in EM density. Our results correspond also with those of Tandon and associates 115, who reported that schizophrenic patients who were drug free only for 2-4 weeks (when the effect of neuroleptics is still present) displayed higher EM density in comparison with schizophrenic patients who were drug free for a longer time or drug naive. Tandon and co-workers 115 did not present data on positive symptoms after drug withdrawal. It is, however, not rare that the withdrawal of neuroleptics causes the exacerbation of psychotic (positive) symptoms 125. Although the relationships between clinical symptoms and both REM sleep variables (REM sleep percentage and EM density) are in the same direction, it is worth mentioning the difference. The increase of REM sleep percentage is determined mainly by the predominance of negative symptoms, while the decrease of EM density is related to the level of positive symptoms. However, this is at least one possible interpretation of both relationships in the frame of the search activity concept. I suggest that the increased REM sleep percentage in group II is caused by the absence of search activity in patients with predominantly negative symptoms. Negative symptoms (apathy, flattened affect, low social activity, inattentiveness, lack of spontaneity in behavior and in mental activity, poverty of speech), like depression, display a lack of search activity that requires REM sleep for its compensation. However, in contrast to depression, negative schizophrenia is not accompanied by the regular decrease of REM sleep latency. It may depend on the different context of the renunciation of search in depression and negative schizophrenia. While in depression, and especially in anxious and agitated depression, renunciation of search is usually combined with increased affective tension, in negative schizophrenia it is usually accompanied by flattened affect. At the same time, any form of schizophrenia may be combined with depressive symptoms, and this can be an additional factor determining the lack of linear relationships between positive/negative symptom ratio and REM sleep. There is another possible interpretation of the normal REM sleep latency in our patients. It has been shown 121,125 that REM latency correlates with SWS time. The exacerbation of psychosis following acute neuroleptic withdrawal may determine the suppression of SWS and as a result the decrease of REM latency 125. At the same time, the relapse of psychosis after neuroleptic withdrawal decreases the total REM time. Thus, the relapse of psychosis may cause a paradoxical combination of decreased REM sleep latency, decreased REM time and decreased phasic REM activity, while stable neuroleptic treatment, like in our investigation, causes a combination of increased REM duration and REM activity with normal REM sleep latency. However, the relationships between positive/ negative symptoms and sleep structure may be even more complicated and contradictory. Not only may the exacerbation of positive symptoms suppress SWS, Ganguli and associates 122 showed also that SWS and negative symptoms were negatively correlated, which may influence REM latency, especially in non-medicated schizophrenic patients. Positive symptoms represent very particular, misdirected and maladaptive, but at the same time very intense, search activity 20. Paranoid ideation and hallucinations correspond to the definition of search activity: active overt or covert behavior without definite forecast. Auditory hallucinations are associated with increased metabolic activity in brain centers for inner speech, i.e. that represent active verbalization. A schizophrenic patient cannot be sure about future 'events' in the artificial world he is building for himself, or about outcomes of his interactions with this world. At the same time, the person remains sensitive to all events and outcomes related to this artificial world. The hyperactivity of the brain catecholamine system in positive symptom schizophrenia also corresponds to the notion of search activity 20. In healthy subjects, EMs in REM correlate with the active participation of the dreamer in dream scenarios and with dream content 35,75. Active events in dreams represent search activity that has to compensate the renunciation of search in the previous wakefulness 5. If search activity during wakefulness is already high, there is no need for high EM density in REM sleep. From my point of view, this may explain how neuroleptic withdrawal precipitated a psychotic relapse associated with the decline in REM sleep duration and EM activity 125, and explains why positive symptoms correlated negatively with EM density in our investigation. The peculiarity of the FNE in schizophrenia also can be explained in the frame of the search activity concept. Neylan and associates 125 did not find FNE in schizophrenic patients on and off haloperidol therapy. In our investigations 10, 127, FNE according to our criteria (see above) was present only in 35% of schizophrenic patients, less than in MD. Interestingly, the age of patients who demonstrated FNE was significantly higher than the mean age of patients without FNE. Positive symptoms were higher in patients with FNE while negative symptoms were almost equal in both groups. When the ratio of positive/ negative symptoms was > 0.6, FNE was present in 71% of all cases, whereas when this ratio was < 0.4, FNE was present only in 17%. It was difficult to discriminate the influence of age and positive symptoms on the difference between REM sleep latency on the first and the second nights. The ratio of the first REM period to the total amount of REM sleep was not increased in the first night of schizophrenic patients, and in patients with FNE this variable was less than on the second night. In patients with a FNE, sleep efficiency on the first night was lower and the number of awakenings higher than on the first night in the control group of healthy subjects who demonstrated a FNE. Thus, it is possible to suggest that there is an exaggerated FNE in schizophrenic patients with relatively high positive symptoms in comparison to normal subjects. Although patients with FNE were, in general, older than patients without FNE, it is very unlikely that FNE is directly determined by age. In the investigation of Ganguli and co-workers 122 almost half of the patients suffering from schizophrenia demonstrated FNE although all patients were young. In normal young subjects, FNE is a typical finding 45, 130. There are some reasons to suggest that positive symptoms play a key role in the FNE: (I) Schizophrenic patients with positive symptoms, like PTSD patients, display an exaggerated orienting reaction 131. At the same time, patients whose negative symptoms predominate demonstrate a failure to respond to the environment 132. (2) Schizophrenic patients with positive symptoms display higher reactivity to sensory and affective stimuli in comparison to patients with negative symptoms 135. Such sensitivity may increase with age. (3) A relatively higher response of the brain monoamine system in patients with positive symptoms, in comparison to patients with negative symptoms 134, may also explain the appearance of the FNE as an exaggerated reaction in a new environment. (4) As we have already shown, positive symptoms decrease REM sleep pressure. As a result, REM sleep pressure does not prevent FNE (the reverse of the situation in depressed patients with high REM sleep pressure and a low REM sleep flexibility). It is interesting to discuss the difference between schizophrenic patients with positive symptoms and the above-mentioned patients with psychotic depression. The latter do not display FNE. It is possible to speculate that hallucinations and delusions of schizophrenic patients cause an active and outside oriented, although inappropriate and misdirected, search behavior while delusions in depressed patients are often mood-congruent and are related to the feeling of guilt, worthlessness, failure, to ruminative self-blaming and self-annihilation. Such delusions provide a basis for renunciation of search. It was shown 135 that psychotic depressive patients, in comparison to melancholic patients and to those with major depression without psychotic features, were significantly more likely to demonstrate marked psychomotor disturbances, to report feeling sinful and guilty, and to suffer from constipation, terminal insomnia, appetite loss and loss of interest and pleasure. Our approach to the difference between psychotic mood-congruent depression and schizophrenia is in line with data 136 that cortisol non-suppression on the dexamethasone suppression test is most prominent in depression with mood-congruent delusions, in comparison to both non-psychotic depression and depression with mood-incongruent delusions. Lower rates of non-suppression were also observed in schizophrenia 137. These data suggest that psychotic features in depression and schizophrenia may have different significance, and this proposition is confirmed by our data derived from sleep investigations. Our group has shown that melatonin treatment of chronic schizophrenia definitely restored FNE in these patients in comparison to placebo treatment 138. There are different possibilities to explain these data. It is possible that melatonin enhances the alertness of schizophrenic patients in the unfamiliar situation as an outcome of the improved sleep quality on melatonin treatment during the previous nights 138. The restoration of FNE may result from the modulation of dopaminergic activity by melatonin in the neuroleptic-treated patients with schizophrenia. Finally, we have investigated the relationship between objective sleep variables and subjective sleep estimation in schizophrenia 139. There are no systematic investigations of this topic. Patients with chronic schizophrenia demonstrated a high, and unexpected, ability to estimate correctly the duration of sleep latency. The correlation between objective and subjective sleep latency in patients was higher than in healthy subjects and in depressed patients. In healthy subjects and patients with mood disorders the duration of wakefulness before sleep onset is probably overestimated, owing to emotional tension caused by sleep delay. This emotional tension may interfere with the ability to estimate sleep latency. In chronic schizophrenic patients with a relative predominance of negative symptoms and with blunted affect, alteration of sleep onset does not cause any emotional reaction and its subjective estimation is not distorted. At the same time, the subjective estimation of the duration of wakefulness in sleep correlated in schizophrenic patients with EM density in REM sleep. The same correlation was found in patients with MD (see above). As stressed previously, in healthy subjects EM density correlates with some features of dream reports. However, in schizophrenic patients we found a negative correlation between EM density and dream reports. It is possible to speculate that schizophrenic patients, as well p as patients with MD, perceive mental activity in REM sleep as wakefulness, and it is an important sign of the REM sleep functional insufficiency. According to the search activity concept, such functional insufficiency of REM sleep may play an important role in the pathogenesis of different mental disorders.
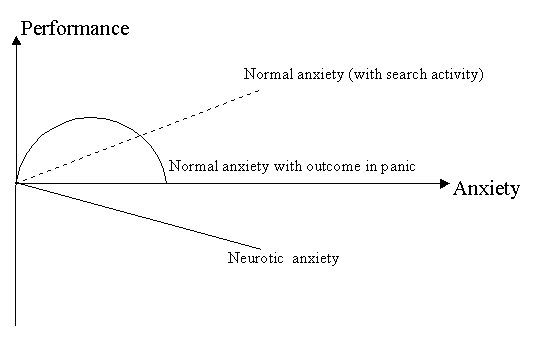
ADAPTIVE VS. MALADAPTIVE EMOTIONAL TENSION (ANXIETY) AND SLEEP It is well known that anxiety is one of the main reasons for sleep disturbances. Anxiety causes prolonged sleep latency, decreased total sleep time, reduced SWS and reduced sleep efficiency partly due to the increased number of awakenings. Beyond the alterations of the objective sleep structure, anxiety may cause a negative subjective estimation of sleep 140. Sleep latency may be overestimated; awakenings filled with anxious feelings might be more easily fixed in memory than those without and will result in reports of poor sleep. Moreover, awakenings in NREM sleep, especially in the first sleep cycles, may cause the underestimation of the depth and duration of the preceding part of sleep 84. The alterations of REM sleep variables in anxiety are less definite than alterations of non-REM sleep or of sleep duration. This may relate to the problem of the definition of anxiety. Emotional tension (anxiety) is not a united psychophysiological state. It can be either maladaptive or adaptive in both nature and outcome. Adaptive emotional tension helps the subject to solve problems and to overcome obstacles, and has no negative outcome on health. Despite the theoretical and practical importance of the distinction between adaptive and maladaptive emotional tension, neither adequate theoretical approaches nor valid methods appear to distinguish between the two in past research. According to general activation theory, an optimal level of emotional tension is adaptive in helping the individual to solve problems and to overcome obstacles without any negative outcome for the organism. The Yerkes-Dodson law postulate that when the level of tension is extremely low or high, its outcome is regarded as negative with respect to performance, adaptation and health 141,142. However, there are many exceptions to the Yerkes-Dodson law. First, pathological emotional tension (neurotic anxiety) is always harmful, with respect not only to health but also to performance, thereby decreasing the effectiveness of the latter in a linear manner 143,144. Second, even a very high level of emotional tension can promote the activity of the subject (Figure 3.1). According to Dienstbier 145, naturally evoked peripheral catecholamines never seem to be too high for optimal performance. In addition, when methods of coping were available, even very high arousal levels failed to elicit discomfort and negative emotions 146. I suggest that the difference between productive emotional tension (the normal anxiety of a healthy individual in a state of stress), instrumental in mobilizing all psychological and physical resources to overcome obstacles, and unproductive emotional tension that hampers successful activity is determined by the presence or absence of search activity in the structure of emotional tension. Until search activity is present, anxiety is adaptive. All forms of maladaptive anxiety (like panic and neurotic anxiety) do not correspond to search activity, both according to the theoretical notion and clinical observations. During panic the results of the activity are not considered at any stage and cannot be used for the correction of behavior. Panic behavior during catastrophic events is usually displaced by or combined with depression 147,148. Panic may represent an exaggeration of normal anxiety that corresponds to the final point of the curvilinear line displaying relationships between emotional tension and adaptation or performance (see Figure 3.1). In this final point the emotional tension is very high while performance and adaptation are low. This happens, in particular, when vital motivations are increased and fear of the consequences of failure predominates over constructive attempts to find a reasonable solution to the problem; such fear does not allow the subject to follow up, to the end, any way of search and to use the current outcome of his/her own activity in order to correct this way. Finally, such fear deteriorates behavior. In panic, like in other states of emotional tension without search activity, the expenditure of brain monoamines exceeds their synthesis, and eventually there comes a pronounced drop in their levels and a secondary depression.
Figure 3.1 The relationships between different types of anxiety and performance
Neurotic anxiety, according to the psychodynamic concept, is a consequence of the repression of an unacceptable motive from consciousness. The repression can be regarded as a purely human variant of renunciation of search, of the modes of realization, of the unacceptable motive in overt behavior, and modes of integration of it with other realized behavioral orientations. Thus, repression simply represents a state of giving up in front of the inner motivational conflict and its outcome is, speaking metaphorically, a mental "freezing". It is the reason why neurotic anxiety caused by repression is maladaptive 85 and does not follow the Yerkes-Dodson law. Benzodiazepines have different outcomes for adaptive and maladaptive emotional tension. It is well known that benzodiazepines sometimes enhance and sometimes diminish the behavior (motor) response to threatening signals 149. In moderate doses, they do not block active avoidance or even increase it, but they usually block freezing as a sign of renunciation of search 150. Maladaptive anxiety is more sensitive to anxiolytics, and because of the competition between renunciation of search and search behavior, freezing can be replaced by active avoidance. However, benzodiazepines also block adaptive anxiety containing search activity, by increasing the sensory threshold for any meaningful information that can cause frustration. Imagine, for instance, a lecturer who is worrying about the possible outcome of his lecture. Even if it is a mobilizing and adaptive emotional tension, subjectively it is accepted as an unpleasant anxiety and has a negative connotation for the person: he prefers not to worry. If in order to cope with this anxiety he uses benzodiazepines, the adaptive mobilization is reduced and the lecture will be less successful. Ledoux and Gorman 151 have recently confirmed our approach to anxiety. They stressed that a shift from passive suffering to active coping is the best way to treat anxiety caused by recent social events. The search activity concept helps to solve some contradictions related to the dynamic of autonomic patterns in different forms of anxiety. Thus, there is a marked difference between the habituation of orienting reactions in healthy subjects versus subjects with psychopathology. In healthy subjects, the amplitude of the electrodermal reaction (EDR) during the first two orienting reactions is higher and the habituation is faster, while the spontaneous electrodermal activity is lower 152. If the spontaneous electrodermal activity reflects an inner emotional tension not relevant to the external task, and the evoked EDR reflects the emotional tension during decision making 153, it can be suggested that pathological subjects are overwhelmed by their emotional tension, which causes hyperarousal. This high arousal, however, reflects maladaptive emotional tension, is in competition with search activity and blocks normal forms of behavior requiring search activity, in particular, the orienting reaction. From my point of view, this is a more reasonable explanation than the explanation according to the 'law of initial level' 154. Wilder proposed that the higher the initial level of activity of the physiological system, the lower the reaction of the system on the relevant task. However, in healthy subjects involved in a meaningful activity and highly motivated, autonomic variables may increase regardless of their initial level 155,156. Wilder's law seems to be relevant only for pathological anxiety. Thus, the low autonomic reaction in pathologically anxious persons may not relate to the high initial level by itself, but rather to the inability of the subject to accept the challenge of the task that requires search activity. Sleep structure is also able to separate adaptive and maladaptive anxiety. We analyzed sleep change in 27 healthy students on a post-examination night in comparison to post-holiday nights 2. Thirty minutes before the examination and 30 min after, we recorded their heart rate frequency, arterial blood pressure and bioelectrical activity of the orbicular muscles of the mouth. The latter, as other autonomic variables, is very sensitive to emotional tension. The control data on these indices were obtained on one of the term days. Before the examination all subjects showed an increase in autonomic variables and muscle tension compared with the control measures. However, after the examination, two groups could be separated. The group I subjects' (16 students) indices dropped to the control level, whereas the indices of subjects in group II remained at a stable high level. Students of group II demonstrated a significant increase in REM sleep percentage on the post-examination night compared with the control night and with the post-examination night of the group I subjects. On the post-examination night, the subjects of group II showed a positive correlation between the total duration of sleep and the proportion of REM sleep (0.74) and a negative correlation (- 0.6) between the duration of REM sleep and that of delta sleep. The subjects of group II showed worse results in resolving logical tasks on the evening after the examination (before sleep) than on the following morning; also, their results were worse than those of group I. They were working relatively faster but they made significantly more mistakes - thus they behaved in a more impulsive way. It is important to note that subjects in both groups did not differ according to the objective outcome of the stressful situation -all students passed the examination. Thus, stable emotional tension was not caused by the negative outcome of the examination. We have proposed a hypothesis that the subjects of group I exhibited normal emotional tension, which ensured psychophysiological mobilization for overcoming the stressful situation and diminished when the situation passed. The subjects of group II displayed maladaptive emotional tension. It did not diminish after the stressful situation was over and required REM sleep for its disappearance. Additional support for this approach comes from data demonstrating that slow post-stress catecholamine decline is accompanied by poor performance, neuroti-cism and pathological anxiety 157. Almost everybody who has ever passed examinations is familiar with such maladaptive tension. It prevents concentration on the particular text the subject is studying because the subject is overwhelmed with fears according to the predicted negative outcome of examinations. Instead of reading the text the subject is continually counting the number of pages he has yet to go. It is possible to suggest that coming examinations, as every stressful condition, provoke in such subjects deep and old neurotic complexes and feelings of inferiority, and this is the reason why the anxiety does not disappear even after the actual stressful situation is over. In order to cope with these activated complexes and feelings that provoke renunciation of search the subject has to go through REM sleep. Maladaptive anxiety determines paradoxical relationships between sleep disturbances and daytime alertness. Recently, we investigated daytime wakefulness/sleepiness and night sleep structure in patients with sleep apnea and in patients with depression/anxiety 158. We found that in patients with sleep apnea the ability to remain awake during soporific circumstances measured by the maintenance of wakefulness test (MWT) correlated negatively with stage 1 sleep. The latter correlated negatively with the total sleep time and with SWS. It looks very natural - the more disturbed the night's sleep due to sleep apnea, the more difficult it is to maintain daytime alertness. However, in patients with depression/anxiety, MWT scores correlated negatively with total sleep time and SWS (stage 3), while the multiple sleep latency test correlated negatively with total sleep time and sleep efficiency. This means that the more disturbed the night's sleep, the less easily is the subject able to go to sleep during the day. Maladaptive tension disturbs nighttime sleep and at the same time prevents daytime sleep. It determines a very well known paradoxical combination of subjective sleepiness (caused by nighttime sleep disturbance) with high irritation and inability to sleep.
ROLE OF SLEEP IN MEMORY FUNCTION: REM SLEEP, BEHAVIOR AND MEMORY The outlined ideas concerning the function of REM sleep and dreams presented in part of this chapter can help to solve many debatable problems of sleep psychophysiology. Among such problems, in particular, is the role of sleep in memory and learning. Jenkins and Dallenbach 159 in their classical investigation demonstrated that recall was diminished less by a night of sleep than by an equivalent amount of wake time. However, in their theory no active role in memory consolidation and storage was ascribed to sleep; it was considered only as a period free from sensory interference. The discovery of REM sleep and different NREM sleep stages elicited many modern investigations on the active role of these sleep stages in memory. The recent review by Stickgold 160 summarized data of investigations performed on animals and humans. It was shown that in animals, REM sleep increases after training on very different but complex tasks such as the multiple-goal maze, shuttle avoidance, classical conditioning and operant bar press, mostly after the so-called unprepared learning according to Seligman 161. This REM sleep increase appeared in special time periods (REM sleep 'windows' 162), often just before the critical level of success in the learning process was achieved 163 and correlated with the successful learning of new material. REM sleep deprivation performed after the learning process in such critical periods ('windows') disturbs retention. At the same time, performance of simple tasks was not impaired by REM sleep deprivation 164. Investigations performed on humans have also shown that the relationship between the learning process and sleep depends on the task peculiarity. Tasks related to declarative memory (word recognition, retention of the word list or paired associates) are REM sleep independent and REM deprivation produced no effect on these tasks 165, while tasks related to procedural memory that require the development of new and flexible perceptual or motor skills - like mirror writing, the word fragment completion task or the Tower of Hanoi task 166 - are REM dependent 160. Declarative memory as opposed to procedural memory depends on SWS 167-169. According to the above-mentioned data, many authors 160,162,164,170,171 suggest that REM sleep per se plays an important role in the consolidation of unusual and emotionally significant information, but not in the retention of ordinary information. However, it was shown that the creative (unusual) task, in most cases, is not solved in REM sleep per se, although after a dream with an active position of the dreaming subject the creative task in subsequent wakeful-ness is solved more successfully 172. On the other hand, REM sleep deprivation does not influence the learning process, if the learning prior to the deprivation was performed with intensity and success, and the task was realized during wakefulness 170. This means that REM sleep is not obligatory for the retention of information even if the task by itself is complex. There are some additional arguments against the idea of the direct participation of REM sleep in consolidation mechanisms: (1) During the active search behavior in the stressful situation, consolidation of the new experience is essential for survival, but under these conditions REM sleep does not increase, and even decreases. (2) The REM sleep percentage increase under neuroleptic treatment 118 has no beneficial effect on memory. (3) Activating drugs, like amphetamine, have a tendency to suppress REM sleep but, at least, do not disturb memory 173. (4) Despite marked suppression of REM sleep, antidepressants on the whole do not disrupt learning/memory 174. As already mentioned, REM sleep becomes longer at certain stages during the solution of emotionally significant complex problems, which the animal or human subject is originally unprepared to handle 170. There is evidence suggesting that the greatest increase of REM sleep occurs during the 24 h before the critical level of success in the learning process 175,176. REM sleep deprivation carried out directly in this period impedes it. It is possible to suggest that REM sleep is particularly important for the animal during the critical period of an acute conversion from the previous stereotypical behavior to the new and flexible style of behavior. It might be a sign that REM sleep plays only an indirect role in the process of retention by carrying out its main function - compensation of renunciation of search and restoration of search activity 4. A complex problem that the subject is not ready to solve, a difficult task that requires unprepared learning, may cause renunciation of search with much greater probability than a simple one, especially at the early stages of the solution when failures prevail over successes. In experiments on animals "the learning situation imposed is undoubtedly the most important and traumatic event in the life of the organism" 171, and such an event has a significant chance of producing a giving-up reaction (renunciation of search). Actually, the non- learning rats (rats that remain below the learning criteria) displayed a considerable majority of freezing responses daring training and thereby received the highest number of electric shocks 177. If search activity in REM sleep is unavailable (due to REM deprivation) and, consequently, does not overcome the state of renunciation of search, the latter itself will make it impossible to find the right solution or keep it in memory. REM deprivation after training leaves memory in a labile form and delays the process of consolidation 178, presumably due to the predominance of the maladaptive state of renunciation. Therefore, the lengthening of REM sleep exactly before the critical point in the learning, after which the animal fully develops the habit, is not surprising. However, in some cases the lengthening of REM sleep may be insufficient to compensate for the very prominent state of renunciation of search. In such cases, REM sleep may increase precisely in those animals that are the least successful in forming the new habit 179, resembling the increase of REM sleep in depression. It is possible to suggest that in non-learning rats the compensatory REM sleep system is usually weak and functionally insufficient. This hypothesis corresponds to data 177 that the average duration of REM sleep episodes of baseline sleep of non-learning rats is significantly shorter in comparison to learning rats. These episodes may be less stable. In such cases stressful conditions of training may cause a relative increase of REM sleep that is nevertheless insufficient for compensation. The peculiarity of neuronal activity in different functional states also does not confirm the theory of REM sleep's direct participation in memory consolidation. According to the dynamic of neuronal activity, in quiet rest and in SWS, information flows from hippocampus to neocortex, while in REM sleep and during active exploration in wakefulness 160,180, information flows in the opposite direction - from neocortex to hippocampus. Thus according to this variable, as well as according to die hippocampal theta-rhythm, active exploration in wakefulness and REM sleep are very similar to each other and opposite to SWS. This confirms our conclusion about active exploration (search activity) in functionally sufficient REM sleep. However, it shows also that it is not necessary to ascribe memory consolidation by itself to REM sleep. The above-mentioned explanation of the indirect contribution of REM sleep in memory function is here more relevant. Renunciation of search makes any functions in wakefulness and sleep, including memory, less efficient, and if REM sleep overcomes renunciation of search and restores search activity it has also a positive outcome on memory. Furthermore, this approach provides an explanation for data being difficult to explain by the concept of memory consolidation in REM sleep. For instance, what is the functional meaning of the REM sleep increase after immobilization stress or learned helplessness? What sort of information is it so necessary to consolidate by means of REM sleep after such an experience? The subject needs only to overcome the unproductive state of renunciation of search elicited by these conditions. This means that the concept of the indirect positive role of REM sleep in memory functions seems to be more comprehensive and broad. This concept also allows explanation of some data showing that REM sleep has a double effect on retention: facilitative and inhibitory 169. Latash and Manov 167 have performed an investigation on the task of declarative memory. Human subjects were awakened in the first or second cycle in delta sleep close to REM sleep or after the first or second episode of REM sleep. Every subject passed all four experimental conditions, but every night each subject had only one awakening, during which he was tested for the amount of retention of material he had learned before going to sleep. The authors found that retention after the second REM sleep episode was lower than immediately before this episode. The addition of a short period of time that corresponds to the second REM sleep period had an unexpected inhibitory influence on the declarative memory retention. At the same time, REM sleep facilitated the positive effect of SWS on retention. From my point of view, this facilitative effect was indirect and can be explained as an outcome of the general positive effect of REM sleep on mental state due to the restoration of search activity. A direct negative effect of REM sleep on retention may be due to the interference between dream images and learned material. This double effect of REM sleep may explain the different outcomes of REM sleep deprivation in different conditions. If the learning process per se causes renunciation of search, the requirement in REM sleep has to be particularly high and REM deprivation will inhibit retention. If the learning process stimulates search activity, REM sleep deprivation may sometimes even facilitate retention. In addition, by accepting this concept, some other contradictions are avoided. As mentioned above, there are data that declarative memory tasks depend on SWS while procedural memory tasks depend on REM sleep. Of course, there are differences between some substantial aspects of declarative and procedural memory tasks - for instance, procedural tasks are more complex and the subject is less prepared for such tasks and this can cause giving up (renunciation of search) during training. However, it is difficult to imagine that these two sorts of task are even opposite according to the essential process of information retention and memory consolidation and that they are related to the opposite directions of flow of information. It was shown recently 181 that several brain areas activated during the execution of a serial reaction time task during wakefulness were significantly more active during REM sleep in subjects previously intensively trained on the task than in non-trained subjects. In addition, this task was performed better after sleep than before sleep. According to the authors, these results support the hypothesis that memory traces are processed during REM sleep in humans. However, the same brain structures may be involved in search activity during training in wakefulness as well as in REM sleep, and these data can be explained without ascribing to REM sleep a direct participation in memory function. On the other hand, it was shown 182 that dreams may represent, in a very direct way, the previous waking experience and problems the subject was preoccupied with in previous wakefulness. If subjects have been involved in task performance (and even periodically failed in this performance) they can imagine this task in dreams and the relevant brain area may be activated during such imagination, as happens during the imagination in wakefulness. Does it really mean that such imagination contributes to memory consolidation? It seems to be very doubtful because usually people do not see in their dreams any elements of information they have to consolidate in their memory. The outcome of REM sleep deprivation before training on memory function deserves a short special discussion. A long REM sleep deprivation on the wooden platform before learning destroys simple active avoidance 185, while a short REM deprivation can even stimulate active avoidance 26. If REM deprivation is short, then search activity is frustrated, but brain monoamines and physiological mechanisms responsible for search activity are not yet exhausted. The change from the frustrating conditions on the wooden platform to the conditions of learning and testing may cause a rebound of motor activity and search activity 4. Such new conditions can even promote some forms of learning, for instance, active avoidance. Conversely, if REM deprivation is prolonged, a renunciation of search activity appears, and the whole process of learning is destroyed. Memory traces then become more sensitive to disturbing influences. Thus, REM sleep deprivation before learning creates distress that can interfere with memory.
REFERENCES 1. Rotenberg VS, Arshavsky W. Search activity and its impact on experimental and clinical pathology. Act Nero Super (Praha) 1979;21:105-15 2. Rotenberg VS, Arshavsky V. REM sleep, stress and search activity. Waking Sleeping 1979;3:235-44 3. Rotenberg VS. Search activity in the context of psychosomatic disturbances, of brain monoamines and REM sleep function. Pavlov J Biol Sci 1984;19:1-15 4. Rotenberg VS. Sleep and memory I. The influence of different sleep stages on memory. Neurosci Biobehav Rev 1992;16:497-502 5. Rotenberg VS. REM sleep and dreams as mechanisms of the recovery of search activity. In MofEtt A, Kramer M, Hoffmann R, eds. The Functions of Dreaming. Albany, NY: State University of New York Press, 1993;261-92 6. Rotenberg VS, Boucsein W. Adaptive vs. maladaptive emotional tension. Genet Soc Gen PsycholMonog 1993; 119:207-32 7. Rotenberg VS, Schattenstein AA. Neurotic and psychosomatic disorders: interdependence in terms of search activity concept. Pavlov J Biol Sci 1990;25:43-7 8. Rotenberg VS, Korosteleva IS. Psychological aspects of the search activity and learned helplessness in psychosomatic patients and healthy testees. Dynamische Psychiatric/ Dynamic Psychiatry 1990;120:1-13 9. Rotenberg VS, Sirota P, Elizur A. Psycho-neuroimmunology: searching for the main deteriorating psychobehavioral factor. Genet Soc Gen Psychol Monogr 1996;122:329-46 10. Rotenberg VS, Hadjez J, Martin T, et al. First night effect in different forms of schizophrenia (pilot investigation). Dynamische Psychiatrie/ Dynamic Psychiatry 1998;172/173:421-30 11. Rotenberg VS. Sleep after immobilization stress and sleep deprivation: common features and theoretical integration. Crit Rev Neurobiol 2000;14:225-31 12. Rotenberg VS. The revised monoamine hypothesis: mechanism of antidepressant treatment in the context of behavior. Int Physiol BehavSci 1994;29:182-8 13. Rotenberg VS. Anorexia nervosa: old contradictions and a new theoretical approach. Int J Psychiatry Clin. Pract 2000;4:89-92 14. Coyne J, Lazarus R. Cognitive style, stress perception and coping. In Kutash I, Schlezinger L, eds. Handbook on Stress and Anxiety: Contemporary Knowledge, Theory and Treatment. San Francisco: Jossey-Bass, 1980:57-79 15. Seligman M. Helplessness. On Depression, Development and Death. San Francisco: W.H. Greeman, 1975 16. Katz R, Roth K, Carrol B. Acute and chronic stress effects on open field activity in the rat: implication for a model of depression. Neurosci Biobehav Rev 1981;5:247-51 17. Lehnert H, Reinstein D, Strowbridge B, Wurtman R. Neurochemical and behavioral consequences of acute uncontrollable stress: effect of dietary tyrosine. Brain Res 1984;303:215-23 18. Kohno Y, Hoaki Y, Glavin G, et al. Regional rat braia-' NA turnover in response to restraint stress. Pharmacol Biochem Behav 1983;19:287-90 19. Paul A, van Dongen G. The human locus coeruleus in neurology and psychiatry. Prog Neurobiol 1981:17:97-139 20. Rotenberg VS. An integrative psychophysio-logical approach to brain hemisphere functions in schizophrenia. Neurosci Biobehav Rev 1994;18:487-95 21. Cohen H, Edelman A, Bower R, Delmont W. Sleep and self-stimulation in the rat. Presented at the Eleventh Annual Meeting of the Associated Professional Sleep Societies (APSS), New York, 1975:abstr75 22. Putkonen P, Putkonen A. Suppression of paradoxical sleep following hypothalamic defense reactions in cats during normal conditions and recovery from PS deprivation. Brain Res 1971:26:334-47 23. Kafi S, Bouras C, Constantinidis J, Gaillard J-M. Paradoxical sleep and brain catecholamines in the rat after single and repeated administration of alpha-methyl-parathyrosine. Brain Res 1977:135:123-34 24. Rotenberg VS. Richness against freedom: two hemisphere functions and the problem of creativity. Eur J High Ability, 1993;4:11-19 25. Gaillard J. Involvement of noradrenaline in wakefulness and paradoxical sleep. In Wauquier A, Gaillard JM, Monti JM, Radulovacki M, eds. Sleep: Neurotransmitters and Neuromodulators. New York: Raven Press, 1985:57-67 26. Onvuii TN,' Lortkipanidze ND. Effect of paradoxical sleep deprivation on the learning and memory. In Oniani TN, ed. Neurophysiology of Motivation, Memory and Sleep-Wakefulness Cycle. Tbilisi, Georgia: Metsniereba, 1985;4:214-34 27. Cartwright R, Luten A, Young M, Mercer P, Bears M. Role of REM sleep and dream affect in overnight mood regulation: a study of normal volunteers. Psychiatry Res 1998;81:1-8 28. Cartwright R, Young MA, Mercer P, Bears M. Role of REM sleep and dream variables in the prediction of remission from depression. Psychiatry Res 1998;80:249-55 29. Kupfer DJ. REM latency - a psychobiological marker for primary depressive disease. Biol Psychiatry 1976; 11:159-74 30. Adrien J, Dugovic C, Martin P. Sleep-wakefulness patterns in the helpless rat. Physiol Behav 1991;49:257-62 31. Rotenberg VS. Learned helplessness and sleep: discussion of contradictions. Homeostasis 1996:37:89-92 32. Kuiken D, Sikora S. The impact of dreams on waking thoughts and feelings. In Moffitt A, Kramer M, Hoffmann R, eds. The Functions of Dreaming. Albany: State University of New York Press, 1993;419-76 33. Morrison A. Central activity states: overview. In Beckman AL, ed. The Neural Basis of Behavior. New York: Spectrum, 1982:3-17 34. Rosenzweig S. A test for types of reaction to frustration. Am J Orthopsychiatry 1935;5:395-403 35. Rotenberg VS. Functional deficiency of REM sleep and its role in the pathogenesis of neurotic and psychosomatic disturbances. Pavlov J Biol Sci 1988:23:1-3 36. Kramer M. The selective mood regulatory function of dreaming: an update and revision. In Moffitt A, Kramer M, Hoffinann R, eds. The Functions of Dreaming Albany, NY: State University of New York Press, 1993:139-96 37. Purcell S, Moffitt A, Hoffinann R. Waking, dreaming and self-regulation. In Moffitt A, Kramer M, Hoffinann R, eds. The Functions of Dreaming. Albany, NY: State University of New York Press, 1993:197-260 38. Greenberg R, Pearlman C. The private language of the dream. In Natterson J, ed. The Dream in Clinical Practice. New York: Aronson, 1980:85-96 39. Rotenberg VS. Word and image: the problem of context. Dynamische Psychiatrie Dynamic Psychiatry 1979;59:494-8 40. Rotenberg VS. Sleep dreams, cerebral dominance and creation. Pavlov J Biol Set 1985;20: 53-8 41. Rotenberg VS, Arshavsky W. Psychophysiology of hemispheric asymmetry: the 'entropy' of right hemisphere activity. Integr Physiol Behav Sci 1991 ;26:183-8 42. Rotenberg VS, Arshavsky W. Right and left brain hemisphere activation in the representatives of two different cultures. Homeostasis 5 1997:38:49-57 43. Agnew H, Webb W, Williams R. The first night effect: an EEC study of sleep. Psychophysiology 1966;2:263-6 44. Sharpley A, Solomon R, Cowen P. Evaluation of first night effect using ambulatory monitoring and automatic sleep stages analysis. Sleep 1988:11:274-6 45. Woodward SH, Bliwise DL, Friedman MJ, Gusman FD. First night effects in post-traumatic stress disorder inpatients. Sleep 1996; 19:312-17 46. Coates T, George J, Killen J, Marchini E, Hamilton S. Thorensen C. First night effect in good sleepers and sleep maintenance insomniacs when recorded at home. Sleep 1981;4:293-8 47. Riemann D, Low H, Shredil M, Wiegand M, Dippel B, Berger M. Investigation of morning and laboratory dream recall and content in depressive treatment with trimipramine. PsychiatrJ Univ Ott 1990; 15:93-9 48. Armitage R, Rochlen A, Fitch Th, Madhukar T, Rush AJ. Dream recall and major depression: a preliminary report. Dreaming 1995:5:189-98 49. Kaminer H, Lavie P. Sleep and dreams in well-adjusted and less adjusted Holocaust survivors. In Stroebe MS, Stroebe W, Hansson RO, eds. Handbook of Bereavement. USA: Cambridge University Press, 1993:331-4 50. Ross RJ, Ball WA, Sullivan KA, Caroff SN. Sleep disturbance as the hallmark of post-traumatic stress disorder. Am J Psychiatry 1989:146:697-707 51. Inman DJ, Silver SM, Doghramji K. Sleep disturbance in post-traumatic stress disorder: a comparison with non-PTSD insomnia./ Traum Stress 1990:3:429-37 52. Rosen J, Reynolds CF, Yeaer AL, Houck PR, Hurwitz LF. Sleep disturbances in survivors of the Nazi holocaust. Am J Psychiatry 1991; 148: 62-6 53. Mellman ThA, Kumar A, Kulick-Bell R, Kumar M, Nolan B. Nocturnal/daytime urine noradrenergic measures and sleep in combat-related PTSD. Biol Psychiatry 1995;38:174-9 54. Mellman ThA, Nolan B, Hebding J, Kulick-Bell R, Dominguez R. A polysomnographic comparison of veterans with combat-related PTSD, depressed men, and non-ill controls.' Sleep 1997:20:46-51 55. Greenberg R, Pearlman C, Campel D. War neuroses and the adaptive function of REM' sleep. Br J Med Psychol 1972:45:27-33 56. Kauffmann CD, Reist C, Djenderedjian A, Nelson JN, Haier RJ. Biological markers of affective disorders and post-traumatic stress disorder: a pilot study with desipramine./ Clin Psychiatry 1987;48:366-7 57. Schlosberg A, Benjamin M. Sleep patterns in three acute combat fatigue cases. J Clin Psychiatry 1978;9:546-9 58. Lavie P, Hefez A, Halperin G, Enoch D. Long-term effects of traumatic war-related events on sleep. Am J Psychiatry 1979;136:175-8 59. Hefez A, Metz L, Lavie P. Long-term effects of extreme situational stress on sleep and dreaming. Am J Psychiatry 1987; 144:344-7 60. Glaubman H, Mikulincer M, Porat A, Wasserman O, Birger M. Sleep of chronic post-traumatic patients. J Traumat Stress 1990;3: 255-63 61. Van Kammen WB, Christiansen C, van Kammen DP, Reynolds CF. Sleep and the prisoner-of-war experience - 40 years later. In Giller IR, ed. Biological Assesment and Treatment of Posttraumatic Stress Disorder. Washington DC: American Psychiatric Press, 1990:159-72 62. Ross RJ, Ball WA, Dinges DF. Rapid eye movement sleep disturbance in posttraumatic stress disorder. Biol Psychiatry 1994;35:195-202 63. Kulka RA, Fairbrank JA, Hough RL, Marmar, CR, Weiss DW. Trauma and the Vietnam Generation: Findings from the National Vietnam Veterans Readjustment Study; New York: Brunner/Mazel, 1990 64. Benca RM, Obermeyer WH, Thisted RA, Gillin C. Sleep and psychiatric disorders. A meta-analysis. Arch Gen Psychiatry 1992;49: 651-68 65. Woodward SH, Friedman MJ, Bliwise DL. Sleep and depression in combat-related PTSD inpatients. Biol Psychiatry 1996;39:182-92 66. Emilien G, Penasse C, Charles G, Martin D, Lasseaux L, Waltregny A. Post-traumatic stress disorder: hypothesis from clinical neuropsychology and psychopharmacology research. MJ Psychiatry Clin Pract 2000;4:3-18 67.JBauer M, Priebe S, Graf KJ, et al. Psychological and endocrine abnormalities hi refugees from East Germany, II. Serum levels of cortisol, prolactin, luteinizing hormone, follicle stimulating hormone and testosterone. Psychiatry Res 1994;51:75-85 68. Carrol BJ, Feinberg M, Greden JF. et al. A specific laboratory test for the diagnosis of melancholia. Arch Gen Psychiatry 1981;38:15-22 69. Whalley LJ, Borthwick N, Copolov D, et al. Glucocorticoid receptors and depression. Br MedJ 1986:292:859-61 70. Yehuda R, Boisoneau D, Lowy MT, Giller FL. Dose response 'changes in plasma cortisol and lymphocyte glucocorticoid receptors following dexamethasone administration in combat veterans with and without posttraumatic stress disorder. Arch Gen Psychiatry 1995;52: 583-93 71. Perry BD, Giller EL, Southwick SM. Altered platelet alpha2-adrenergic binding sites in posttraumatic stress disorder. Am J Psychiatry 1987:144:1511-12 72. Gershuni BS.,Thayer JF. Relations among psychological trauma, dissociative phenomena, and trauma-related distress: a review and integration. Clin Psychol Rev 1999;19:631-57 73. Rotenberg VS. Right hemisphere insufficiency and illness in the context of search activity concept. Dynamische Psychiatrie /Dynamic Psychiatry 1995:150/151:54-63 74. Schore AN. The effects of early relational trauma on right brain development, affect regulation, and infant mental health. Infant Ment Health J 2001;22:201-69 75. Hong Ch Ch-h, Potkin SG, Antrobus JS, Dow BM, Callaghan GM, Gillin JCh. REM sleep eye movement counts correlate with visual imagery in dreaming: a pilot study. Psychophysiology 1997:14:377-81 76. Kinzie JD, Fredrickson RH, Ben R, Fleck J, Karls W. Posttraumatic stress disorder among survivors of Cambodian concentration camps. Am J Psychiatry 1984:141:645-9 77. Goldstein G, van Kammen W, Shelly C, Miller D, van Kammen D. Survivors of imprisonment in the Pacific Theatre during World War II. Am J Psychiatry 1987:144:1210-14 78. Kramer M, Schoen LS, Kinney L. Nightmares in Vietnam veterans. / Am Acad Psychoanal 1987:15:67-81 79. Husain AM, Miller PP, Carwile ST. REM sleep behavior disorder: potential relationship to post-traumatic stress disorder. J Clin Neurophysiol 2001:18:148-57 80. Greenberg R, Pillard R, Pearlman Ch. The effect of dream (stage REM) deprivation on adaptation to stress. Psychosom Med 1972;34: 257-62 81. Kramer M, Schoen LS, Kinney L. The dream experience in dream-disturbed Vietnam veterans. In van den Kolk BA, ed. Post-traumatic Stress Disorder: Psychological and Biological Sequelae. Washington DC: American Psychiatric Press, 1984:81-95 82. Lavie P, Kaminer H. Dreams that poison sleep: dreaming in Holocaust survivors. Dreaming 1991; 1: 11-21 83. Bleich A, Attias Y, Dagan Y, Lavie P, Shalev A, Lerrer B. Psycho-neuro-physiologic characteristics of combat veterans with posttraumatic stress disorder. Clin Neuropharmacol 1990; 13(Suppl 2):334 84. Rotenberg VS. Sensitivity, neuroticism and sleep disturbances: some controversial problems. Waking Sleeping 1980:4:271-9 85. Rotenberg VS, Michailov AN. Characteristics of psychological defense mechanisms in healthy testees and in patients with somatic disorders. Homeostasis 1993;34:54-8 86. PlutchikR,KellermanH, ConteHR. A structural theory of EGO defenses and emotions. In Izard E, ed. Emotions in Personality and Psychopathology. New York: Plenum Publishing, 1979:229-57 87. Grieser C, Greenberg R, Harrison R. The adaptive function of sleep: the differential effects of sleep and dreaming on recall. J Abnormal Psychol 1972:80:280-6 88. Kupfer DJ, Frank E, Grochorinski VJ, Gregor M, McEacharon AB. Electroencephalographic sleep profiles in recurrent depression. Arch Gen Psychiatry 1998;45:678-81 89. Ansseau M, Kupfer DJ, Reynolds CF III. Internight variability of REM latency in major depression: implications for the use of REM latency as a biological correlate. Biol Psychiatry 1985:20:489-505 90. Reynolds CF III, Kupfer D. Sleep in depression. In Williams RZ, Karacan I, Moore CA, eds. Sleep Disorders, Diagnosis and Treatment. New York: John Wiley, 1988:147-64 91. Benson E, Zarcone VP. Rapid eye movement sleep eye movements in schizophrenia and depression. Arch Gen Psychiatry 1993;50:474-82 92. Riemann D, Berger M. EEC sleep in depression and in remission and the REM sleep response to the cholinergic agonist RS 86. Neuropsychopharmacology 1989:2:145-52 93. Thase ME, Reynolds CF III, Frank E, el al. Polysomnographic studies of unmedicated depressed men before and after cognitive behavior therapy. Am Psychiatry 1994;151: 1615-22 94. Rush AJ, Erman MK, Giles DE, et al. Polysomnographic findings in recently drug-free and clinically remitted depressed patients. Arch Gen Psychiatry 1986:43:878-84 95. Buysse DJ, Kupfer DJ, Frank E, Monk TH, Ritenour A. Electroencephalographic sleep studies in depressed outpatients treated with interpersonal psychotherapy. II. Longitudinal studies at baseline and recovery. Psychiatry Res 1992:40:27-40 96. Kupfer DJ, Ulrich RF, Coble PA, et al. The application of automated REM and slow wave sleep analysis (normal and depressives). Psychiatr Res 1984:13:325-34 97. Reynols CF, Kupfer DJ. Sleep research in affective illness: state of the art circa 1987. Sleep 1987:10:199-215 98. Vogel GW, Vogel F, McAbur RS, Thurmond AJ. Improvement of depression by REM sleep deprivation. Arch Gen Psychiatry 1980:37:247-53 99. Wirz-Justice A, Van den Hoofdaker RH. Sleep deprivation in depression: what do we know, where do we go? Biol Psychiatry 1999;46: 445-53 100. Greenberg R, Pearlman Ch. Cutting die REM sleep nerve: an approach to the adaptive role of REM sleep. Perspect Biol Med 1974;17: 513-21 101. Rotenberg VS, Shamir E, Barak Y, Indursky P, Kayumov L, Mark M. REM sleep latency and wakefulness in the first sleep cycle as markers of major depression. A controlled study vs. schizophrenia and normal controls. Prog Neuro-Psychopharmacol Biol Psychiatry 2002;26: 1211-15 102. Rupfer DJ, Frank E, Ehlers CL. EEC sleep in young depressives: first and second nights effects. Biol Psychiatry 1989;25:87-97 103. Reynolds CF III, Newton TF, Shaw DM, Coble PA, Kupfer DJ. Electroencephalographic sleep findings in depressed outpatients. Psychiatry Res 1982;6:65-72 104. Coble P, Foster C, Kupfer D. Electroence-phalographic sleep diagnosis of primary depression. Arch Gen Psychiatry 1976;33:1124-7 105. Akiskal HS, Lemmi H, Yerevanian B, King D, Belluomoni J. The utility of the REM latency test in psychiatric diagnosis: a study of 81 depressed outpatients. Psychiatry Res 1982;7: 101-10 106. Rotenberg VS, Hadjez J, Kimhi R, et al. First night effect in depression: new data and a new approach. Biol Psychiatry 1997;42:267-74 107. Rotenberg VS, Kayumov L, Indursky P, et al. REM sleep in depressed patients: different attempts to achieve adaptation. J Psychosom Res 1997;42:565-75 108. Reynolds CF III, Hoch CC, Buysse DJ, et al. Sleep in late-life recurrent depression: changes during early continuation dierapy widi nor-triptyline. Neuropsychopharmacology 1991;5: 85-96 109. Rotenberg V Kayumov L, Indursky P, et al. Slow wave sleep redistribution and REM sleep eye movement density in depression: towards die adaptive function of REM sleep. Homeostasis 1999;39:81-9 110. Indursky P, Rotenberg VS. Change of mood during sleep and REM sleep variables. Int J Psychiatry Clin Pract 1998;2:47-51 111. Cartwright RD, Lloyd SR. Early REM sleep: a compensatory change in depression? Psychiatry Res 1994;51:245-52 112. Kramer M, Roth T. A comparison of dream content in dream reports of schizophrenic and depressive patients groups. Cvnipr Psychiatry 1973; 14:325-9 113. Rotenberg VS, Indursky P, Kayumov L, Sirota P, Melamed Y. The relationship between subjective sleep estimation and objective sleep variables in depressed patients. Int J Psychophysiol 2000;37:291-7 114. Keshavan MS, Reynolds GF, Kupfer D. Electroencephalographic sleep in schizophrenia: a critical review. Compr Psychiatry 1990;30: 34-47 115. Tandon R, Smrpley JE, Taylor S, et al. Electroencephalographic sleep abnormalities in schizophrenia: relations to positive/ negative symptoms, and previous neuroleptic treatment. Arch Gen Psychiatry 1992;49: 185-94 116. Hudson JI, Lipinski JP, Keck PE, et al. Pblysomnographic characteristics of schizophrenia in comparison with mania and depression. Biol Psychiatry 1993;34:191-3 117. Lauer FChJ, Schreiber W, Pollmacher Th, Holsboer F, Krieg J-Ch. Sleep in schizophrenia: a polysomnographic study of drug-naive patients. Neuropsychopharmacology 1997; 16: 51-60 118. Mendelson WB, Gillin JCh, Wyatt RJ. Human Sleep and its Disorders. New York: Plenum Press, 1977 119. Hiatt JF, Floyd TC, Katz PH, Feinberg I. Further evidence of abnormal non-rapid-eye-movement sleep in schizophrenia. Arch Gen Psychiatry 1985;42:797-802 120. Benson E, Zarcone VP. Low REM latency in schizophrenia. Sleep Res 1985, 14:124 121. Zarcone VP, Benson KL, Berger PA. Abnormal rapid eye movement latencies in schizophrenia. Arch Gen Psychiatry 1987;44:45-8 122. Ganguli R, Reynolds ChF III, Kupfer DJ. Electroencephalographic sleep in young, never-medicated schizophrenics. Arch Gen Psychiatry 1987;44:36-44 123. Kempenaers C, Kerkhofs M, Linkowski P, Mendlewicz J. Sleep EEG variables in young schizophrenic and depressive patients. Biol Psychiatry 1988;24:833-8 124. Riemann D, Gann H, Fleckenstein P, Hohagen F, Olbrich R, Berger M. Effect of RS86 on REM latency in schizophrenia. Psychiatry Res 1991;38:89-92 125. Neylan ThC, van Kammen DP, Kelley ME, Peters JL. Sleep in schizophrenic patients on and off haloperidol therapy. Clinically stable vs. relapsed patients. Arch Gen Psychiatry 1992; 49:643-9 126. Kupfer DJ, Wyatt RJ, Scott J, Snyder F. Sleep disturbance in acute schizophrenic patients. Am J Psychiatry 1970;126:1213-23 127. Rotenberg VS, Hadjez J, Shamir E, Martin T, Indursky P, Barak Y. Sleep structure as a mirror of die clinical state in schizophrenic patients. Dynamische Psychiatrie /Dynamic Psychiatry, 1999;176/179:334-40 128. Kay SR, Fiszbein A, Opler LA. The positive and negative syndrome scale (PANSS) for schizophrenia. Schizophr Bull 1987:13:261-76 ; 129. Foster FG, Kupfer D, Coble P, McParland RJ. Rapid eye movement sleep density. Arch Gen Psychiatry 1976;33:119-23 130. Toussaint M, Luthringer R, Schaltenbrandt N, et al. First night effect in normal subjects and psychiatric inpatients. Sleep 1995; 18: 463-9 131. Kim DK, Shin YM, Kim ChE, Cho HS, Kim YS. Electrodermal responsiveness, clinical variables and brain imaging in male chronic schizophrenics. Biol Psychiatry 1993:33: 786-93 132. Frith CD. The Cognitive Neuropsychology of Schizophrenia. Hove, UK: Lawrence Erlbaum, 1992 133. Docherty NM. Affective reactivity of symptoms as a process discriminator in schizophrenia. J Nerv Ment Dis 1996; 184:535-41 134. Wolkin A, Sanfilipo M, Duncan E, et al. Blunted change in cerebral glucose utilization after haloperidol treatment in schizophrenic patients with prominent negative symptoms. Am J Psychiatry 1996;153:346-54 135. Parker G, Hedzi-Pavlovik D, Brodati H, et al. Sub-typing depression. II. Clinical distinction of psychotic depression and non-psychotic melancholia. Psychol Med 1995;25:825-32 136. Ayuso-Gutierrez JL, Almoguera MI, Carcia-Camba E, Frias JQ, Cabranes JA. The dexamethasone suppression test in delusional depression: further findings. J Affect Disord 1985;8:147-51 137. Schatzberger AF, Rothshild AJ, Langlais PJ, Bird ED, Cole JO. A corticosteroid/dopamine hypothesis for psychotic depression and related states. Psychiatr Res 1985;19:57-64 138. Shamir E, Rotenberg VS, Laudon M, Zisapel N, Elizur A. First-night effect of melatonin treatment in patients with chronic schizophrenia. J Clin Psychopharmacol 2000;20: 691-4 139. Rotenberg VS, Indursky P, Kimhi R, et al. The relationship between objective sleep variables and subjective sleep estimation in schizophrenia. Int J Psychiatry Clin Pract 2000;4: 63-7 140. Ware JC. Sleep and anxiety. In Williams RL, Karacan I, Moore CA, eds. Sleep Disorders. Diagnosis and Treatment. New York: John Wiley & Sons, 1988;189-214 141. Yerkes RM, Dodson JD. The relation of strength of stimulus to rapidity of habit formation. J Comp Neuml Psychol 1908;18:459-82 142. Lens W, De Voider M. Achievement motivation and intelligent test scores: a test of the Yerkes-Dodson hypothesis. Psychol Belg 1980: 49-59 143. Spielberger CD. The effects of manifest anxiety on the academic achievement of college students. Ment Hyg 1962;46:420-6 144. Lader M. The Psychophysiology of Mental Illness. London: Routledge, 1975 145. Dienstbier RA. Arousal and physiological toughness: implications for mental and physical health. Psychol Rev 1989;96:93-104 146. Gal R, Lazarus RS. The role of activity in anticipating and confronting stressful situations./ Hum Stress 1975;1:4-20 147. Zuckerman M. Sensation seeking: a comparative approach to human traits. Behav Brain Sci 1984;7:413-34 148. Rosenbaum DL, Seligman MEP. Abnormal Psychology. New York: Norton, 1989 149. Gray J. The neuropsychology of anxiety: an inquiry into the functions of septohippocam-pal system. Oxford: Oxford University Press, 1982 150. Soubrie P Inferring anxiety and anti-anxiety effects in animals. Behav Brain Sci 1982:5:502-3 151. Ledoux JE, German JM. A call to action: overcoming anxiety through active coping. Am J Psychiatry 2001;158:1953-5 152. Lader M. The orienting reflex in anxiety and schizophrenia. In Kimmel HD, van Olst EH, Orlebeke JF, eds. The Orienting Reflex in Humans. Hilsdale, NY: Erlbaum, 1979: 607-17 153. Rotenberg VS, Vedenyapin AB. GSR as a reflection of decision-making under conditions of delay. Pavlov J Biol Sci 1985;20:11-14 154. Wilder L. Das Ausgangswert-Gesetz' - ein unbeachtetes biologische Gesetz: Seine Bedeutung fur Forschung und Praxis. Klin Wiss 1931;41:1889-93 155. O'Gorman JG, Jamieson RD. Short latency acceleration of human heart rate as a function of stimuli intensity. Percept Motor Skills 1977;45:579-83 156. Lovallo WR, Pincomb GA, Wilson MF. Predicting response to a reaction time task. Heart rate reactivity compared with Type A behavior. Psychophysiology 1986:23:648-56 157. Lader M. Anxiety and depression. In Gale A, Edwards JA, eds. Physiological Correlates of Human Behavior. Individual Differences and Psychopathology. London: Academic Press, 1983;155-67 158. Kayumov L, Rotenberg VS, Buttoo K, Audi Ch, Pandi-Perumal SR, Shapiro CM. Interrelationships between nocturnal sleep, daytime alertness, and sleepiness: two types of alertness proposed. J Neuropsychiatry Clin Neurosci 2000; 12:86-90 , 159 Jenkins JG, Dallenbach KM. Obliviscence during sleep and waking. Am J Psychol 1924; 35:605-12 160. Stickgold R. Sleep: off-line memory reprocessing. Trends Cogn Sci 1998;2:484-92 161. Seligman MEP. On the generality of the laws of learning. Psychol Rev 1970;77:406-18 162. Smith C. Sleep states and learning. A review of the animal literature. Neurosci Biobehav Rev 1985;9:157-68 163. Smith C, Kitahama K, Valatx JL, Jouvet M. Increases in paradoxical sleep in mice during acquisition of a shock avoidance task. Brain Res 1974;77:221-30 164. Pearlman Ch. REM sleep and information processing: evidence from animal studies. Neurosci Biobehav Rev 1979;3:57-68 165. Conway J, Smith C. REM sleep and learning in humans: a sensitivity to specific types of learning tasks. Presented at the 12th Congress of the European Sleep Research Society, Florence, Italy, June 1994 166. Smith C. Sleep states and memory processes. Behav Brain Res 1995;69:137-45 167. Latash LP, Manov GA. The relationship between delta-sleep and REM sleep phasic components with the retention and reproduction of the verbal material learned before sleep. Fiziol Cheloveka 1975;1:262-70 168. Plihal W, Born J. Effects of early and late nocturnal sleep on declarative and procedural memory.J Cognit Neurosci 1997;9:534-47 169. Rotenberg VS. Sleep and memory. II. Investigations on humans. Neurosci Biobehav Rev 1992;16:503-5 170. McGrath MJ, Cohen JB. REM sleep facilitation of adaptive making behavior: a review of the literature. Psychol Bull 1978:85:24-57 171. Smith C. REM sleep and learning: some recent findings. In Moffitt A, Kramer M, Hoffmann R, eds. The Functions of Dreaming. Albany, NY: State University of New York Press, 1993:341-62 172. Montangero J. Dream, problem solving and creativity. In Cavallero C, Foulkes D, eds. Dreaming as Cognition. Harvester Wheatsheaf, 1993:93-113 173. Coenen AML, van Luijtelaar FLJM. Effects of benzodiazepines, sleep and sleep deprivation on vigilance and memory. Acta Neural Belg 1997:97:123-9 174. Vertes RP, Eastman KE. The case against memory consolidation in REM sleep. Behav Brain Sci 2000:23:867-76 175. Leconte P, Hennevin E, Block V. Analyse des effects d'un apprentisage et de son niveau d'acquisition sur le sommeil paradoxal consecutive. Brain Res 1973;49:367-79 176. Pagel I, Pegram V, Vaughan S, Donaldson P, Bridge W. The relationship of REM sleep with learning and memory in mice. Behav Biol 1973:9:383-8 177. Giuditta A, Ambrosini MV^ Montagriese P, et al. The sequential hypodiesis of the function of sleep. Behav Brain Res 1995:69:157-66 178. Fishbein W. The case against memory consolidation in REM sleep: Balderdash! Behav Brain Sci 2000:23:934-6 179. Fishbein W, Kastaniotis C, Chattman I. Paradoxical sleep: prolonged augmentation following learning. Brain Res, 1974;79:61-75t 180. Chrobak JJ, Buzsaki G. Selective activation of deep layer (V-VI) retrohippocampal cortical neurons during hippocampal sharp waves in the behaving rat. J Neurosci 1994;14:1660-70 181. Maquet P, Laureys S, Peignen Ph, et al. Experience-dependent changes in cerebral activation during human REM sleep. Nat Neurosci 2000;3:831-6 182. Greenberg R, Katz H, Schwartz W, Pearlman Ch. A research-based reconsideration of the psychoanalytic theory of dreaming. / Am Psychoanal Assoc 1992:40:531-50 183. Stern WC. Acquisition impairments following rapid eye movements sleep deprivation in rats. Physiol Behav 1971;7:345-52
|