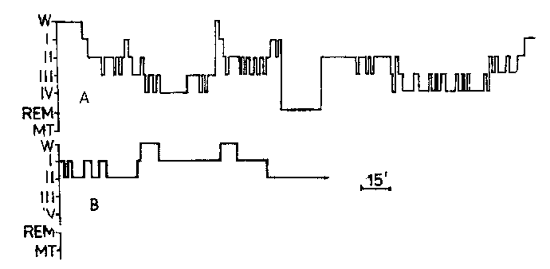
THE STRUCTURE OF NIGHT SLEEP AND CARDIAC RHYTHM IN PATIENTS SUFFERING FOR MANY YEARS FROM HYPO- AND AKINESIAVADIM S. ROTENBERG vadir@post.tau.ac.il, V. I. KOBRIN The structure of night sleep and cardiac rhythm in patients suffering for many years from hypo- and akinesia. V. S. Rotenberg, V. I. Kobrin. Activ. nerv. sup. (Praha) 27, 3, 1985. Polysomnography was performed in 3 patients suffering from long-term akinesia and artificial lung ventilation due to a lesion of the CNS and peripheral nervous system. The cyclic organization of sleep and its transition from stage to stage, as a whole corresponded with those in healthy subjects. However, there was a general tendency in all patients toward a reduction of sleep spindles. There was also a tendency to the stability of heart rate during night sleep. The relationship between deltasleep and sleep spindles and motor activity is discussed. Many questions pertaining to the organization and functional significance of individual sleep stages remain unclear. Hence, the study of the influence of motor activity on sleep structure, on the dynamics of autonomic indices in sleep and in particular, on the dynamics of cardiac rhythm is of certain interest. The object of the present study was the sleep structure and cardiac rhythm in patients who remain long in a state of complete or partial immobility (owing to the paralysis of the musculature of the trunk, limbs and respiratory muscles) and consequently require artificial pulmonary ventilation (APV), for ensuring an adequate metabolism. The authors' attention to this form of pathology stemmed from the fact that in a normal state night sleep is observed after waking, which includes a more or less expressed motor activity, whereas sleep exhibits body movements which influence its structure, in particular, furthering transition to its more superficial stages. Furthermore, the stability of respiration makes it possible to raise some questions regarding the correlation of a number of autonomic indices in sleep. MATERIAL AND FINDINGS Three patients hospitalized at the Institute of Neurology of the USSR Academy of Medical Sciences (in the department headed by Professor L. M. Popova) were examined. Their night sleep was studied by using polysomnography after one adaptation night. The analysis of the sleep structure followed the classification of Rechtschaffen & Kales,1968. Due to the uniqueness of each case the present article sets forth the data of examination of each patient. Patient S., 53. A state after thrombosis of the left vertebral artery, occlusion of the right one, vast infarction in the oral sections of the pons varolii, spastic triplegia and seven-year-old pseudobulbar paresis. Within the same period occurred the loss of voluntary regulation of breathing and of complex behavioural respiratory movements. The patient preserved automatic breathing (metabolic and reflex regulation) and unaided breathing. The duration of the different sleep stages is presented in Table 1 and the structure of the night sleep in Fig. 1. Table 1. Duration of different phases and stages of sleep (in minutes)
On the first night the slow-wave sleep was represented sufficiently
widely showing an even distribution in the early sleep hours but
only one complete cycle (the sleep structure appears to suggest
that the second episode of REM, sleep was replaced by awakening,
which not infrequently becomes revealed by the first examination in
the process of adaptation to the conditions of recording).
Fig. 1. Structure of night sleep of patient S. On ordinate axis
sleep stages.
It is apparent that with the exception of the reduction of the total sleep time and the appropriate diminution of the second stage and REM sleep, the sleep on the patient's first night showed no differences from normal. The second night exhibited a deficit of delta sleep and an increase of the proportion of the first episode of REM sleep, as in the cases of neurotic anxiety or depression. The body movements, as in healthy individuals, concentrated around the episodes of REM sleep and in the waking period in the middle of the night as well. Patient V., 26. At the age of 17 she contracted the generalized form of malignant myasthenia, developing a weakness of the trunk musculature, limbs and respiratory muscles as well as swallowing and speech disturbances. In addition to all these symptoms the excision of the thymus gland added respiratory insufficiency. Since then the patient had been exposed to continuous endotracheal APV by various respirators (Angstrom, Gulber, RO-5). In the past four years the APV had been administered in the conditions of tracheal hermetization by the DP-8 apparatus. Bulbar disturbances (aphagia, anarthria, aphonia) and the absence of her own breathing have been steadily persisting until the present time. Arm and leg movements are limited, and the patient expresses herself only in writing. At the time of examination she had been exposed to continuous APV for a total of eight years and one month. Bronchopulmonary pathology was never observed during the disease.
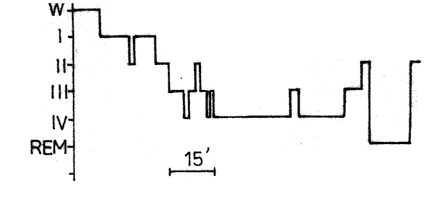
Fig. 2. Structure of night sleep on the second adaptation night of
patient V.
On the first adaptation night the patient displayed a very anxious reaction to the examination and was fearful of damaging the APV system. As a result, she could not fall asleep and therefore the following night had to be regarded as another adaptation night. In its course sleep was recorded polygraphically until the end of the first cycle. This cycle on the second adaptation night was not distinguishable in practically any index from that of healthy testees of the same age: a large proportion of the cycle was taken up by delta sleep. The first episode of REM sleep lasted 10 minutes and slight tendency towards the reduction of the latent period of REM sleep could be noticed, especially if one were to consider that it was virtually a post-deprivation sleep (Fig. 2). In view of this circumstance, one could have expected a long duration of the first cycle.
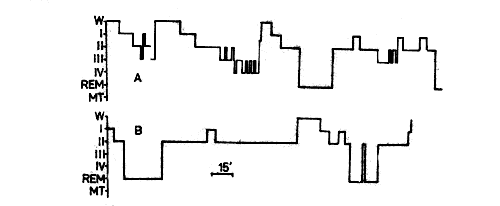
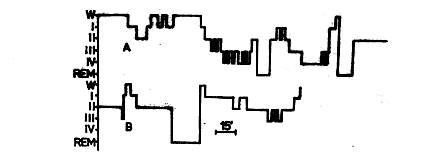
Fig. 3. Structure of night sleep of patient V. On the following night (Fig. 3) the sleep structure in the majority of indices corresponded to that of healthy individuals: the night sleep consisted of three cycles practically equal in time (with the exception of the waking period, when the toilet of the tracheobronchial tree was administered) each of which ended in equally long REM sleep episodes. However, delta sleep (34 minutes) was essentially reduced compared to the age norm. This leads the authors to believe that the high percentage of delta-sleep in the first cycle on the adaptation night reflects the post-deprivation ,rebound" effect. The duration of individual sleep stages is represented in Table 1. Patient P., 39. A case of amyotrophic lateral sclerosis since 1956. Muscular pareses, beginning with the left arm, in the next eight years spread to the musculature of the arms, legs, trunk and respiratory muscle. Aphagia, anarthria and aphonia resulted. Since 1964 APV had been administered. By the time of examination he had had APV administered for 13 years. No pathological processes in the lungs were observed. The patient was fed by the gavage method. Now he exhibits a state of complete paralysis, all his muscles being motionless and athrophic. Retaining the eyeball movements, he uses them in communicating with the personnel with the aid of the ABC. Fig. 4. Structure of night sleep of patient P.
Fig. 5. Histogram of distribution of cardiointervals in waking and
different stages of sleep of patient P. On ordinate axis
proportion of each of the occuring cardiointervals in per cent. On
abscissa axis duration of cardiointervals in seconds. In all indices, including in sleep time (four hours), the two nights were identical. For the duration of the different sleep stages see Table 1. As in the previous patients, the sleep dynamics from one stage to the next are indistinguishable from that of the healthy testees (Fig. 4). Common to all examined patients were some specific features of individual sleep stages. The number of rapid eye movements in their REM sleep did not exceed 34 a minute. In non-REM sleep two patients showed a characteristic combination of alpha- and delta-sleep. Table 2. Cardiointervals in sleep
All patients also had in common a drop in the number of sleep spindles at the second stage. The preserved sleep spindles were frequently low-amplitude and short ones, requiring less than 0.5 second of the recording. With a view to clarifying the dynamics of cardiac rhythm in each cycle at all sleep stages the authors estimated the average pulse frequency and its variations in randomly chosen 100 successive cardiointervals with the subsequent plotting of histograms. The data obtained are summarized in Table 2 and Fig. 5. The authors did not find any essential changes of the cardiac rhythm during sleep but disclosed its stability during the transition from one sleep stage to another and an abrupt narrowing of the range of rhythm variability. The pulse frequency was found to be stable, not only during the transition from one stage to the next within one cycle, but also throughout the night, i.e., the absence of a circadian rhythm, characteristic of healthy individuals, was established. DISCUSSION OF FINDINGS The data presented make it possible to discuss several questions. Above all, attention is attracted by the fact that, as a whole, the patients retained the main symptoms of the cyclic sleep organization. Thus, the many-year absence or considerable decrease of motor activity both in waking and in sleep does not lead to basic changes in the sleep pattern. Consequently, the role of motor activity in the organization of the sleep structure is neither leading nor even essential. All examined patients to varying degrees showed delta-sleep and judging from the results of examinations of patient V., preserved the rebound effect of delta sleep against the background of the general sleep deprivation. There are assumptions as to the major role of the processes which take place during delta sleep in the tissue metabolism, which further the restoration of the tissue, primarily muscular, after exhausting physical loads or, on the contrary, after prolonged hypokinesia (Oswald, 1970; Ryback and Lewis, 1972). The data obtained by the authors cannot be explained from this point of view. In patient P. delta-sleep takes a longer time than in patient V. although patient P. exhibits complete akinesia and muscular atrophy and limited limb movements are preserved in patient V. At the same time, in patient S. there are no expressed atrophies such as in patient P. and he has preserved movements in the right arm and neck and the ability to change his position in bed his delta-sleep being represented maximally. Consequently, there is no direct relationship between delta-sleep, hypokinesia and muscular atrophy. Pronounced delta-sleep can be observed both in the case of maximal and minimal kipokinesia. It can rather be assumed that it depends on the intensity of influx of sensory material and of impressions which in patients S. and P. are greater than in patient V. Patient S. does a great deal of reading, patient P. reads having the pages turned over for him and frequently watches TV whereas patient V. is deprived of both reading and TV watching because even the least strain on her eyes puts a strain on her oculomotor muscles. Thus, the results of these unique examinations make the authors object to the attribution to delta-sleep of a function connected with the restoration of metabolism in the tissues of the entire body. There is more reason to believe that information processes are at work during delta-sleep. This accords with the data of L. M. Mukhametov et al. (Mukhamentov, Supin, Kovalzon, Tumskoi and Tsibulski, 1979; Mukhametov, Supin and Polyakova, 1977), who have demonstrated that delta-sleep deprivation in one hemisphere of a dolphin is accompanied by its compensatory rebound in precisely the same hemisphere. In other words, delta-sleep is necessary above all to the brain, and not to the body as a whole. The present authors, along with others (Jovanovic, 1969; Rotenberg, 1980) have demonstrated that in healthy individuals movements performed in sleep play a certain role in the transitions from the deeper sleep stages to the more superficial ones the role of movements in such transitions increasing in delta-sleep. True, even in neurotic patients, and still more, in healthy individuals, the sleep dynamics from one stage to the next only partially depends on the motor activity: the majority of the transitions to more superficial sleep stages take place without preceding movements. he findings of the present study confirm this conclusion since even patient P., with the complete absence of movements in sleep, exhibits activation shifts in all sleep stages, especially in delta-sleep. Of the qualitative features of sleep of the greatest interest is a drop in the number, amplitude and duration of sleep spindles. It was shown (Vein, Golubev, and Yakhno, 1979) that a similar change of spin activity is observed in sufferers from parkinsonism, especially in the case of its akinetic-rigid form, whereas in the case of the Ziehen-Oppenheim disease the number, amplitude and duration of sleep spindles are increased (Wein and Golubev, 1979).
In the present study the reduction of the sleep spindles was
observed in patients with distinctly expressed muscular
hypotension, both of a central (affection of the frontal horns of
the spinal cord in the case of amyotrophic lateral sclerosis) and
of a peripheral nature (myasthenia). The fact that similar changes
of Stage 2 are also observable in the case of parkinsonism has led
the present authors to assume that the basis of changes such as
these in hypokinesia, common to all their patients. An indirect
confirmation of this idea is the data pertaining to the reduction
of Stage 2 and a drop in the number of the sleep spindles in the
case of prolonged artificial hypokinesia (Ponomaryova, 1976). In
these subjects the general representation of the second sleep stage
was also reduced. Possibly, there is a close connection between the
motor system that ensures motor activity and the stage of sleep
spindles.
As follows from Table 2 and Fig. 5, during REM sleep the frequency and variability of pulse in the examined patients remain unchanged although healthy individuals have been found to exhibit a change (increase and arrhythmia) of heart rate in REM sleep. In two patients (with amyotrophic lateral sclerosis and myasthenia) the possibility to realize the influences of psychic activity on the cardiac rhythm through the vago-sympathetic mechanisms of regulation is preserved but the paralysis of the respiratory musculature makes the influence of psychic activity on the respiration rate impossible. Therefore it is not ruled out that in the normal state, too, psychic activity during REM sleep influences the cardiac rhythm indirectly, through the respiratory system. Motor activity plays a certain role in the formation of shifts in the cardiac rhythm during sleep (Hauri, 1968; Townsend, Johnson et al., 1975). The limitation of this activity in healthy individuals diminishes the variability of cardiac rhythm (Ioffe, 1971). Therefore the abrupt narrowing of the range of variation of cardio-intervals can be directly connected with the long-lasting adynamia. Hence it is worth noting the difference in the frequency and variation of the cardiac rhythm in waking and sleep. The higher variability of heart rate in waking can be due to the changing psychic state in the course of contact with the outside world. If this is the case, psychic factors influence the heart rate dynamics during sleep and waking in different ways and through different mechanisms. The authors are profoundly indebted to Professor L. M. Popova for the possibility to observe the aforementioned patients and the use of relevant clinical material. REFERENCES
Hauri, P.: Effects of Evening Activity on Early Night Sleep.
Psychophysiology 4,
1968, 3, 267277.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||